

Evolučná genetika









Výhody služby
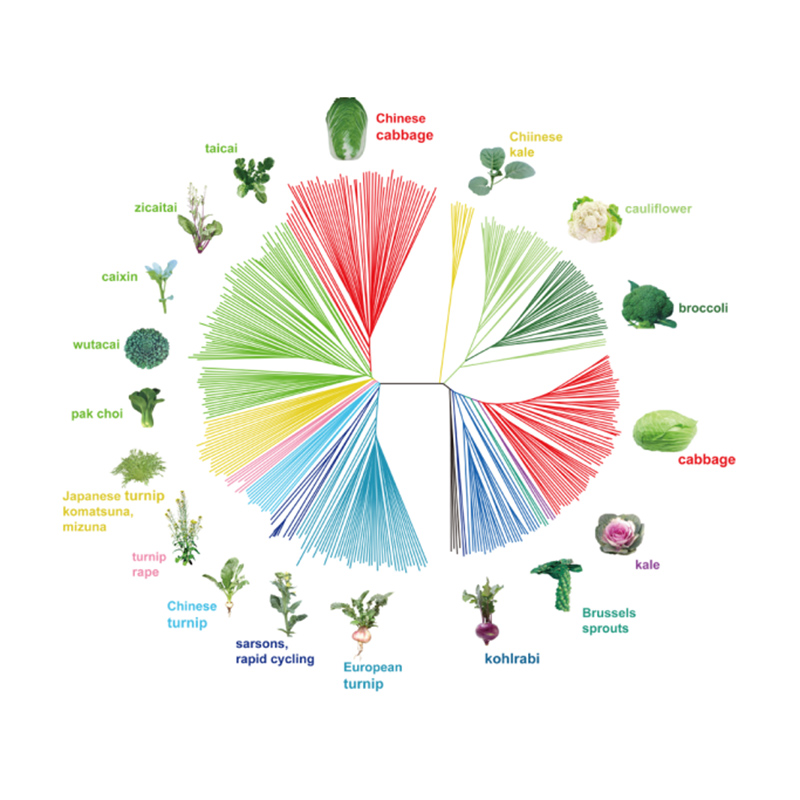
Takagi a spol.,Rastlinný denník, 2013
● Odhad času a rýchlosti divergencie druhov na základe variácií na úrovni nukleotidov a aminokyselín
● Odhalenie spoľahlivejšieho fylogenetického vzťahu medzi druhmi s minimalizovaným vplyvom konvergentnej evolúcie a paralelnej evolúcie
● Vytváranie väzieb medzi genetickými zmenami a fenotypmi s cieľom odhaliť gény súvisiace s vlastnosťami
● Odhad genetickej diverzity, ktorá odráža evolučný potenciál druhov
● Rýchlejší čas obratu
● Rozsiahle skúsenosti: BMK nazbieral rozsiahle skúsenosti s populačnými a evolučnými projektmi za viac ako 12 rokov, ktoré pokrývajú stovky druhov atď., a podieľal sa na viac ako 80 projektoch na vysokej úrovni publikovaných v Nature Communications, Molecular Plants, Plant Biotechnology Journal atď.
Špecifikácie služby
Materiály:
Bežne sa odporúčajú aspoň tri podpopulácie (napr. poddruhy alebo kmene).Každá subpopulácia by mala obsahovať najmenej 10 jedincov (Rastliny >15, pre vzácne druhy sa môže znížiť).
Stratégia sekvenovania:
* WGS možno použiť pre druhy s vysokokvalitným referenčným genómom, zatiaľ čo SLAF-Seq je použiteľný pre druhy s alebo bez referenčného genómu alebo referenčný genóm nízkej kvality.
| Aplikovateľné na veľkosť genómu | WGS | Štítky SLAF (×10 000) |
| ≤ 500 Mb | 10×/jednotlivec | WGS sa odporúča viac |
| 500 Mb – 1 Gb | 10 | |
| 1 Gb – 2 Gb | 20 | |
| ≥2 Gb | 30 |
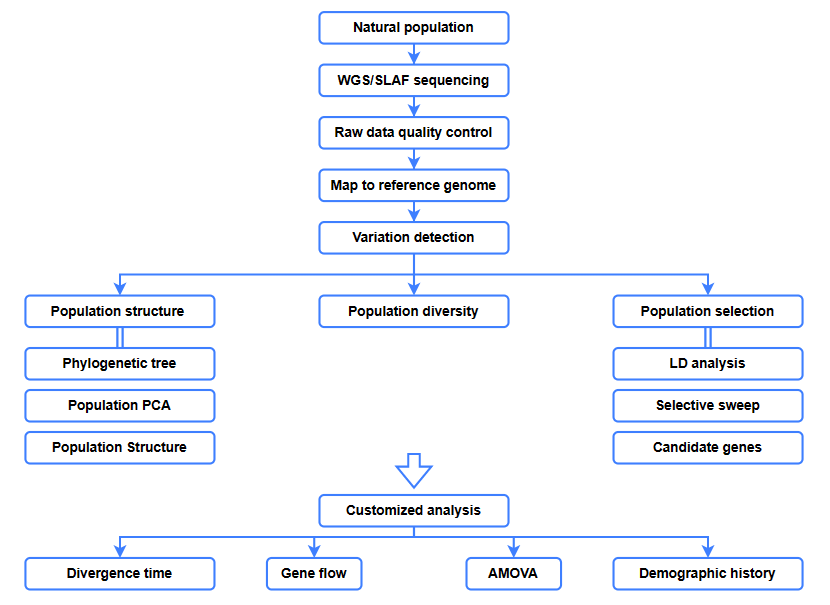
Bioinformatické analýzy
● Evolučná analýza
● Selektívne zametanie
● Tok génov
● Demografická história
● Čas divergencie
Vzorové požiadavky a doručenie
Vzorové požiadavky:
| Druhy | Tkanivo | WGS-NGS | SLAF |
| Zviera
| Viscerálne tkanivo |
0,5 až 1 g
|
0,5 g
|
| Svalové tkanivo | |||
| Cicavčia krv | 1,5 ml
| 1,5 ml
| |
| Hydina/rybia krv | |||
| Rastlina
| Čerstvý list | 1 ~ 2 g | 0,5 až 1 g |
| Okvetný lístok/Stonka | |||
| Koreň/Semeno | |||
| Bunky | Kultivovaná bunka |
| gDNA | Koncentrácia | Suma (ug) | OD260/OD280 |
| SLAF | ≥35 | ≥1,6 | 1,6-2,5 |
| WGS-NGS | ≥1 | ≥0,1 | - |
Tok servisných prác

Dizajn experimentu
Doručenie vzorky
Výstavba knižnice
Sekvenovanie

Analýza dát
Popredajné služby
*Tu zobrazené demo výsledky sú všetky z genómov publikovaných s BMKGENE
1.Evolučná analýza obsahuje konštrukciu fylogenetického stromu, populačnej štruktúry a PCA na základe genetických variácií.
Fylogenetický strom predstavuje taxonomické a evolučné vzťahy medzi druhmi so spoločným predkom.
PCA má za cieľ vizualizovať blízkosť medzi subpopuláciami.
Štruktúra populácie ukazuje prítomnosť geneticky odlišnej subpopulácie z hľadiska frekvencií alel.
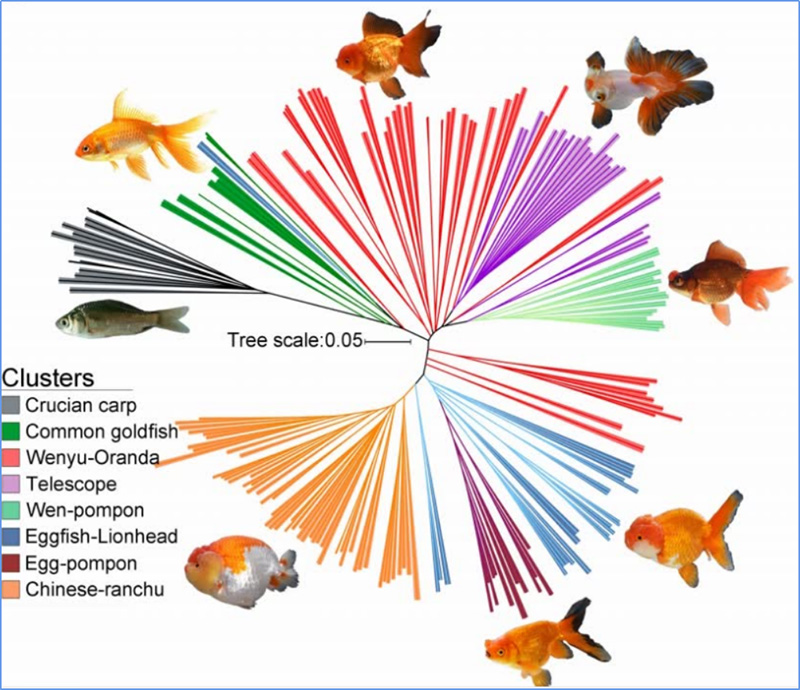
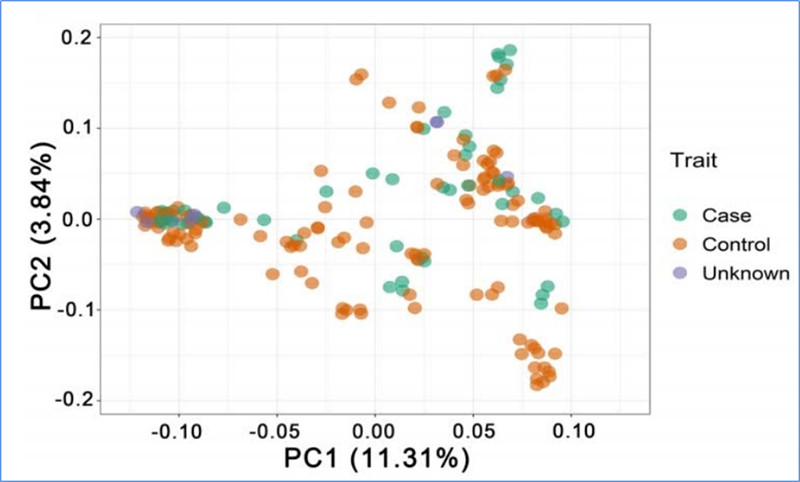
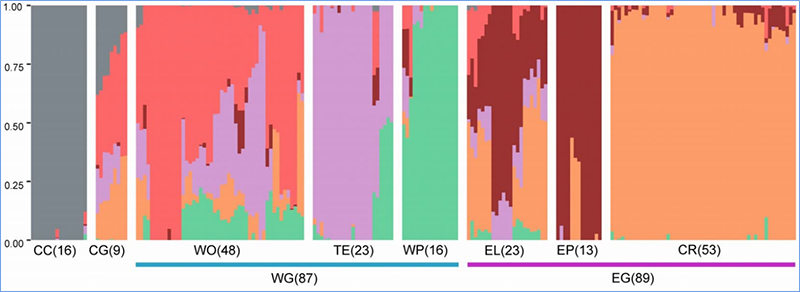
Chen a kol.spol.,PNAS, 2020
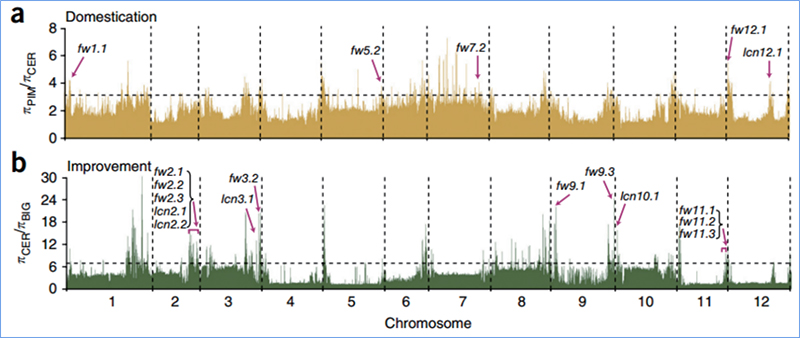
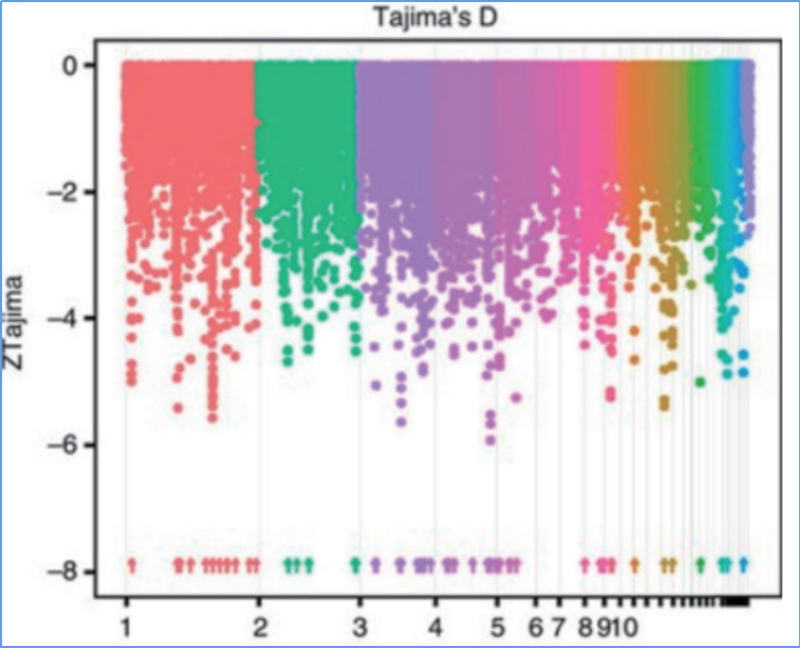
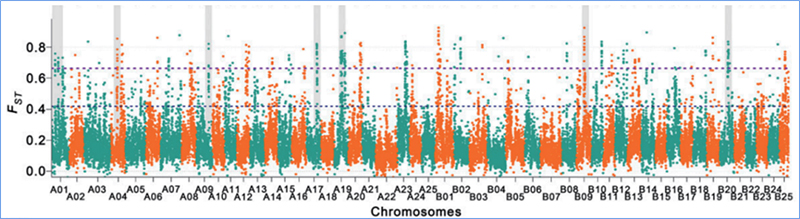
2.Selektívne zametanie
Selektívne zametanie sa týka procesu, ktorým sa vyberie výhodné miesto a frekvencia spojených neutrálnych miest sa zvýši a frekvencia neprepojených miest sa zníži, čo vedie k zníženiu regionálnych.
Detekcia v celom genóme na selektívnych zametacích oblastiach je spracovaná výpočtom populačného genetického indexu (π,Fst, Tajima's D) všetkých SNP v rámci posuvného okna (100 Kb) v určitom kroku (10 Kb).
Nukleotidová diverzita (π)
Tajima D
Fixačný index (Fst)
Wu, et.spol.,Molekulárna rastlina, 2018
3. Tok génov
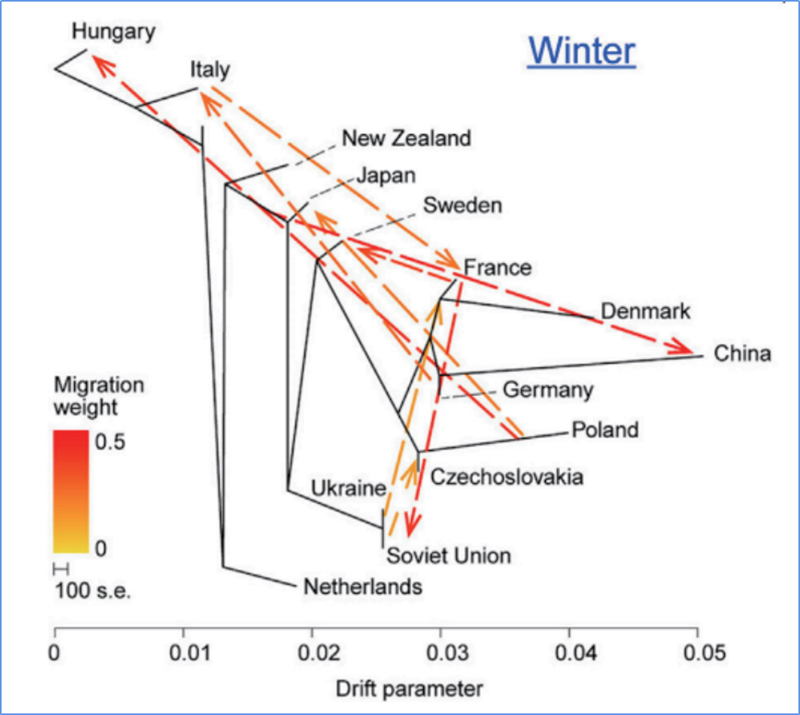
Wu, et.spol.,Molekulárna rastlina, 2018
4.Demografická história
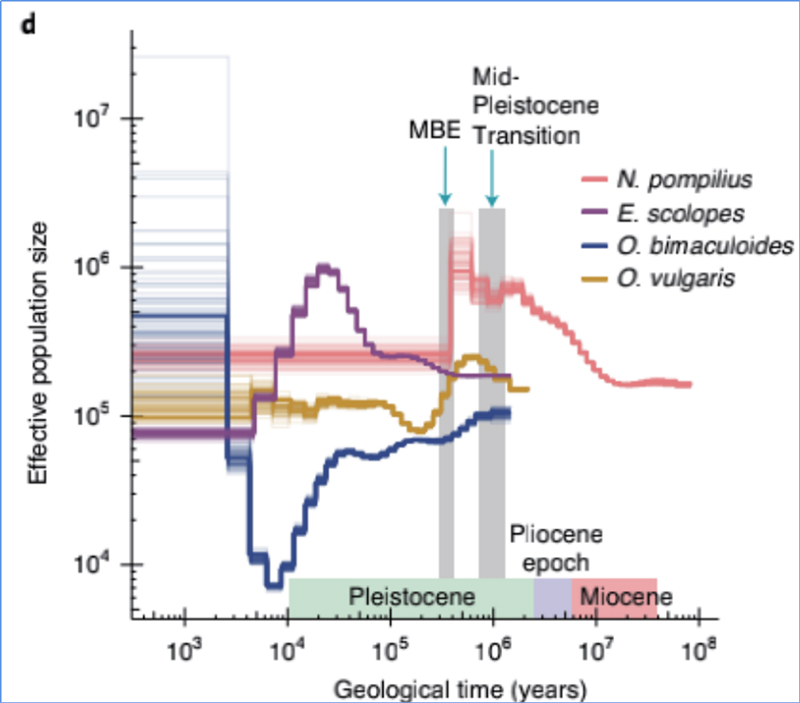
Zhang, et.spol.,Príroda Ekológia a Evolúcia, 2021
5. Čas divergencie
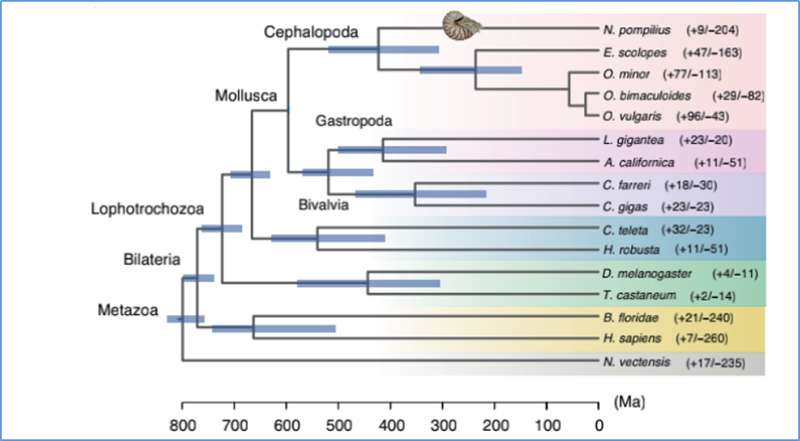
Zhang, et.spol.,Príroda Ekológia a Evolúcia, 2021
Prípad BMK
Mapa genómových variácií poskytuje pohľad na genetický základ výberu jarnej čínskej kapusty (Brassica rapa ssp. Pekinensis)
Publikovaný: Molekulárna rastlina, 2018
Stratégia sekvenovania:
Opätovné sekvenovanie: hĺbka sekvenovania: 10×
Kľúčové výsledky
V tejto štúdii sa spracovalo 194 čínskych kapust na opätovné sekvenovanie s priemernou hĺbkou 10×, čo poskytlo 1 208 499 SNP a 416 070 InDels.Fylogenetická analýza týchto 194 línií ukázala, že tieto línie možno rozdeliť do troch ekotypov, jarný, letný a jesenný.Okrem toho populačná štruktúra a analýza PCA ukázali, že jarná čínska kapusta pochádza z jesennej kapusty v Shandongu v Číne.Tieto boli následne zavlečené do Kórey a Japonska, skrížené s miestnymi líniami a niektoré neskoré odrody z nich boli privezené späť do Číny a nakoniec sa z nich stala jarná čínska kapusta.
Skenovanie celého genómu jarnej čínskej kapusty a jesennej kapusty pri selekcii odhalilo 23 genómových lokusov, ktoré prešli silnou selekciou, z ktorých dva sa prekrývali s kontrolnou oblasťou času skrutkovania na základe mapovania QTL.Zistilo sa, že tieto dve oblasti obsahujú kľúčové gény, ktoré regulujú kvitnutie, BrVIN3.1 a BrFLC1.Štúdiou transkriptómu a transgénnymi experimentmi sa ďalej potvrdilo, že tieto dva gény sa podieľajú na čase skrutkovania.
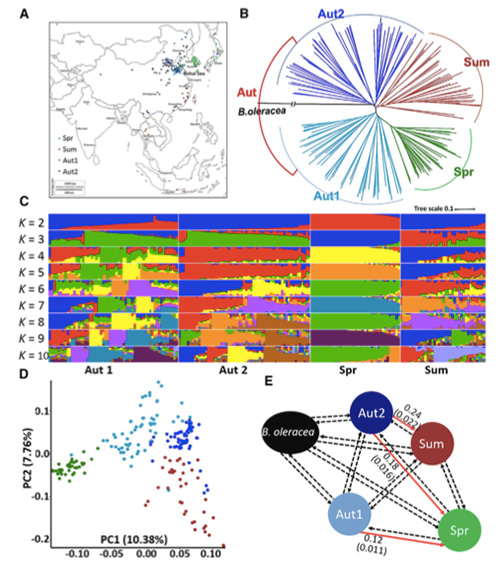 Analýza štruktúry populácie čínskej kapusty | 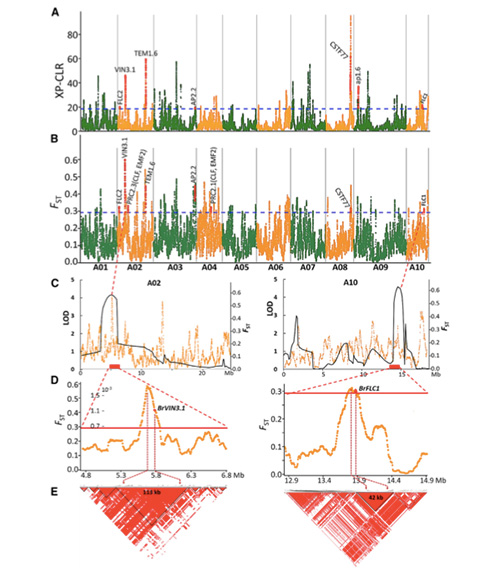 Genetické informácie o výbere čínskej kapusty |
Tongbing a kol."Mapa genómových variácií poskytuje pohľad na genetický základ výberu jarnej čínskej kapusty (Brassica rapa ssp.pekinensis)."Molekulárne rastliny,11(2018):1360-1376.





