

Specific-Locus Amplified Fragment Sequencing (SLAF-Seq)





Tjenestedetaljer
Teknisk ordning
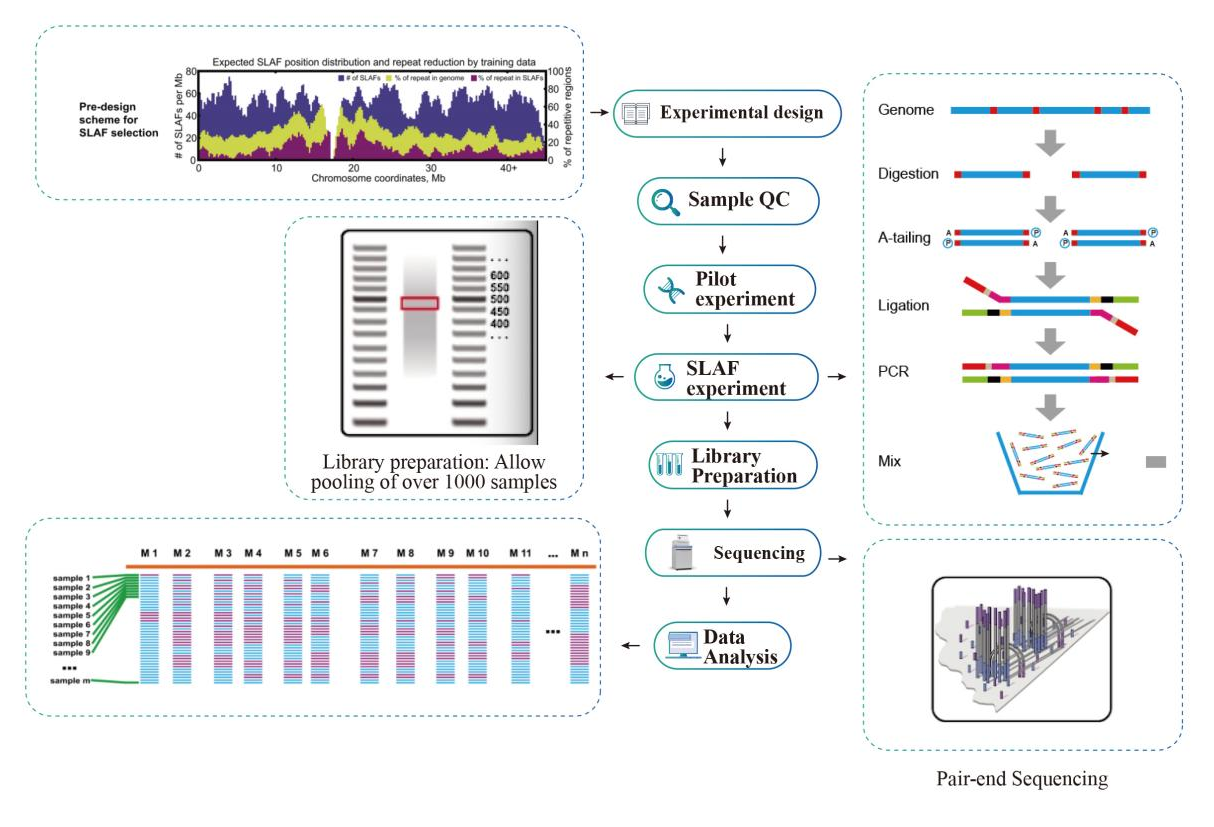
Arbeidsflyt

Tjenestefordeler
Høy effektivitet for oppdagelse av markører- High-throughput sekvenseringsteknologi hjelper SLAF-Seq med å oppdage hundretusenvis av tagger innenfor hele genomet.
Lav avhengighet av genomet– Det kan brukes på arter enten med eller uten referansegenom.
Fleksibel ordningsdesign- Enkelt-enzym, dual-enzym, multi-enzym fordøyelse og ulike typer enzymer, alle kan velges for å imøtekomme ulike forskningsmål eller arter.Forhåndsevaluering i silico brukes for å sikre et optimalt enzymdesign.
Effektiv enzymatisk fordøyelse– Det ble gjennomført forhåndseksperiment for å optimalisere forholdene, noe som gjør det formelle eksperimentet stabilt og pålitelig.Fragmentsamlingseffektivitet kan oppnå over 95 %.
Jevnt fordelte SLAF-brikker- SLAF-tagger er jevnt fordelt i alle kromosomer i størst grad, og oppnår et gjennomsnitt på 1 SLAF per 4 kb.
Effektiv unngåelse av gjentakelser- Repeterende sekvens i SLAF-Seq-data reduseres til lavere enn 5 %, spesielt i arter med høyt nivå av gjentakelser, som hvete, mais, etc.
Omfattende erfaring- Over 2000 lukkede SLAF-Seq-prosjekter på hundrevis av arter som dekker planter, pattedyr, fugler, insekter, vannorganismer, etc.
Egenutviklet bioinformatisk arbeidsflyt- En integrert bioinformatisk arbeidsflyt for SLAF-Seq ble utviklet av BMKGENE for å sikre pålitelighet og nøyaktighet av sluttresultatet.
Tjenestespesifikasjoner
| Plattform | Kons.(ng/gl) | Total (ug) | OD260/280 |
| Illumina NovaSeq | >35 | >1.6(Bind>15μl) | 1,6-2,5 |
Anbefalt sekvenseringsstrategi
Sekvenseringsdybde: 10X/Tag
| Genomstørrelse | Anbefalte SLAF-tagger |
| < 500 Mb | 100K eller WGS |
| 500 Mb - 1 Gb | 100 K |
| 1 Gb -2 Gb | 200 K |
| Gigantiske eller komplekse genomer | 300–400K |
| applikasjoner
| Anbefalt Befolkningsskala
| Sekvenseringsstrategi og dybde
| |
| Dybde
| Tag nummer
| ||
| GWAS
| Prøvenummer ≥ 200
| 10X
|
I følge genomstørrelse
|
| Genetisk evolusjon
| Individer av hver undergruppe ≥ 10; totalt antall prøver ≥ 30
| 10X
| |
Anbefalt prøvelevering
Beholder: 2 ml sentrifugerør
For de fleste prøvene anbefaler vi å ikke konservere i etanol.
Prøvemerking: Prøver må være tydelig merket og identiske med innsendt prøveinformasjonsskjema.
Forsendelse: Tørris: Prøver må først pakkes i poser og begraves i tørris.
Tjenestearbeidsflyt


Prøve QC
Piloteksperiment
SLAF-eksperiment
Bibliotekforberedelse
Sekvensering
Dataanalyse
Ettersalgstjenester
1. Statistikk over kartresultat

2. SLAF markørutvikling

3. Variasjonsmerknad

| År | Tidsskrift | IF | Tittel | applikasjoner |
| 2022 | Naturkommunikasjon | 17.694 | Genomisk grunnlag for giga-kromosomene og giga-genomet til trepion Paeonia ostii | SLAF-GWAS |
| 2015 | Ny fytolog | 7.433 | Domestiseringsfotavtrykk forankrer genomiske regioner av agronomisk betydning i soyabønner | SLAF-GWAS |
| 2022 | Journal of Advanced Research | 12.822 | Genomomfattende kunstige introgresjoner av Gossypium barbadense i G. hirsutum avslører overlegne loki for samtidig forbedring av bomullsfiberkvalitet og -utbytte egenskaper | SLAF-Evolusjonær genetikk |
| 2019 | Molekylær plante | 10,81 | Populasjonsgenomisk analyse og De Novo Assembly avslører opprinnelsen til Weedy Ris som et evolusjonært spill | SLAF-Evolusjonær genetikk |
| 2019 | Naturgenetikk | 31.616 | Genomsekvens og genetisk mangfold av vanlig karpe, Cyprinus carpio | SLAF-Linkage kart |
| 2014 | Naturgenetikk | 25.455 | Genomet til dyrket peanøtt gir innsikt i belgfruktkaryotyper, polyploide evolusjon og domestisering av avlinger. | SLAF-Linkage kart |
| 2022 | Plantebioteknologisk tidsskrift | 9.803 | Identifikasjon av ST1 avslører et utvalg som involverer haiking av frømorfologi og oljeinnhold under soyabønnedomestisering | SLAF-Marker utvikling |
| 2022 | International Journal of Molecular Sciences | 6,208 | Identifikasjon og DNA-markørutvikling for en Wheat-Leymus mollis 2Ns (2D) Disomisk kromosomerstatning | SLAF-Marker utvikling |





