

Evolutionaire genetica









Servicevoordelen
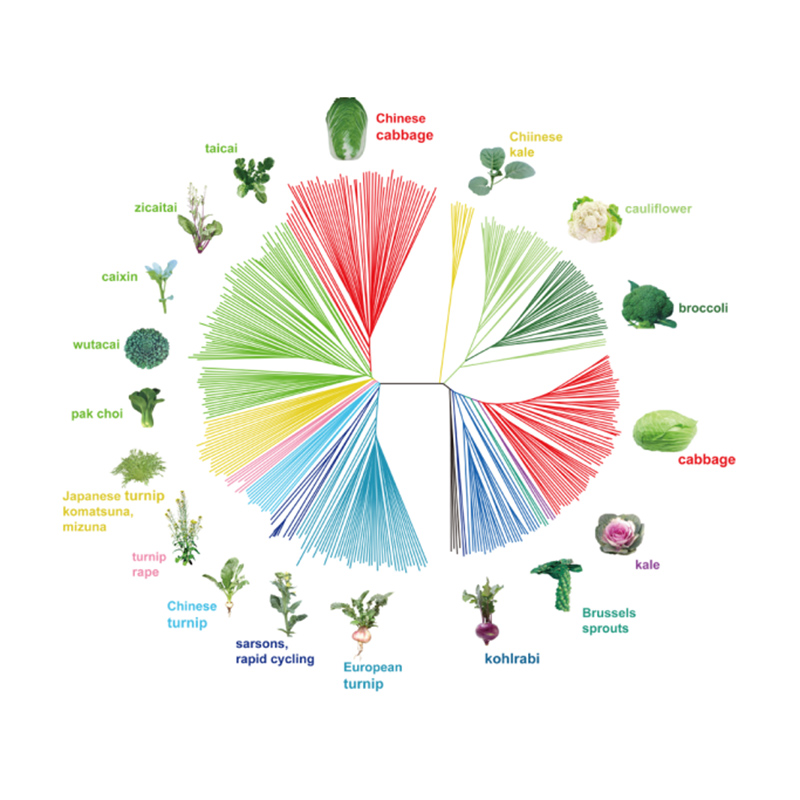
Takagi et al.,Het plantendagboek, 2013
● Het schatten van de tijd en snelheid van de divergentie van soorten op basis van variaties op nucleotide- en aminozuurniveau
● Onthulling van betrouwbaardere fylogenetische relaties tussen soorten met minimale invloed van convergente evolutie en parallelle evolutie
● Het construeren van verbanden tussen genetische veranderingen en fenotypes om eigenschapgerelateerde genen bloot te leggen
● Het schatten van de genetische diversiteit, die het evolutionaire potentieel van soorten weerspiegelt
● Snellere doorlooptijd
● Uitgebreide ervaring: BMK heeft gedurende meer dan 12 jaar een enorme ervaring opgebouwd in populatie- en evolutiegerelateerde projecten, die honderden soorten omvatten, enz. en heeft bijgedragen aan meer dan 80 projecten op hoog niveau die zijn gepubliceerd in Nature Communications, Molecular Plants, Plant Biotechnology Journal, enz.
Servicespecificaties
Materialen:
Normaal gesproken worden ten minste drie subpopulaties (bijvoorbeeld ondersoorten of stammen) aanbevolen.Elke subpopulatie moet niet minder dan 10 individuen bevatten (planten >15, kan worden verminderd voor zeldzame soorten).
Sequentiestrategie:
* WGS kan worden gebruikt voor soorten met een referentiegenoom van hoge kwaliteit, terwijl SLAF-Seq toepasbaar is op soorten met of zonder referentiegenoom, of op referentiegenoom van slechte kwaliteit.
| Van toepassing op genoomgrootte | WGS | SLAF-tags (×10.000) |
| ≤ 500 Mb | 10×/individueel | WGS wordt meer aanbevolen |
| 500 Mb - 1 Gb | 10 | |
| 1 GB - 2 GB | 20 | |
| ≥2 GB | 30 |
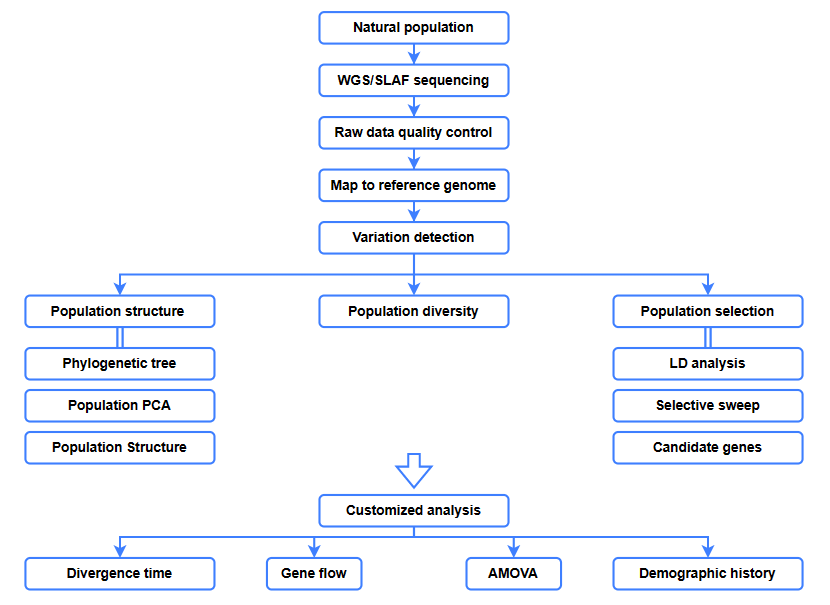
Bio-informatica analyses
● Evolutionaire analyse
● Selectieve sweep
● Genenstroom
● Demografische geschiedenis
● Divergentietijd
Monstervereisten en levering
Voorbeeldvereisten:
| Soort | Zakdoek | WGS-NGS | SLAF |
| Dier
| Visceraal weefsel |
0,5~1g
|
0,5 g
|
| Spierweefsel | |||
| Zoogdierenbloed | 1,5 ml
| 1,5 ml
| |
| Gevogelte/visbloed | |||
| Plant
| Vers blad | 1~2g | 0,5~1g |
| Bloemblaadje/stengel | |||
| Wortel/Zad | |||
| Cellen | Gekweekte cel |
| gDNA | Concentratie | Hoeveelheid (ug) | OD260/OD280 |
| SLAF | ≥35 | ≥1,6 | 1,6-2,5 |
| WGS-NGS | ≥1 | ≥0,1 | - |
Servicewerkstroom

Experimentontwerp
Levering van monsters
Bouw van bibliotheek
Volgorde aanbrengen in

Gegevensanalyse
After-sales diensten
*De hier getoonde demoresultaten zijn allemaal afkomstig van genomen gepubliceerd met BMKGENE
1.Evolutieanalyse omvat de constructie van fylogenetische boom, populatiestructuur en PCA op basis van genetische variaties.
Fylogenetische boom vertegenwoordigt taxonomische en evolutionaire relaties tussen soorten met een gemeenschappelijke voorouder.
PCA heeft tot doel de nabijheid tussen subpopulaties te visualiseren.
De populatiestructuur toont de aanwezigheid van genetisch verschillende subpopulaties in termen van allelfrequenties.
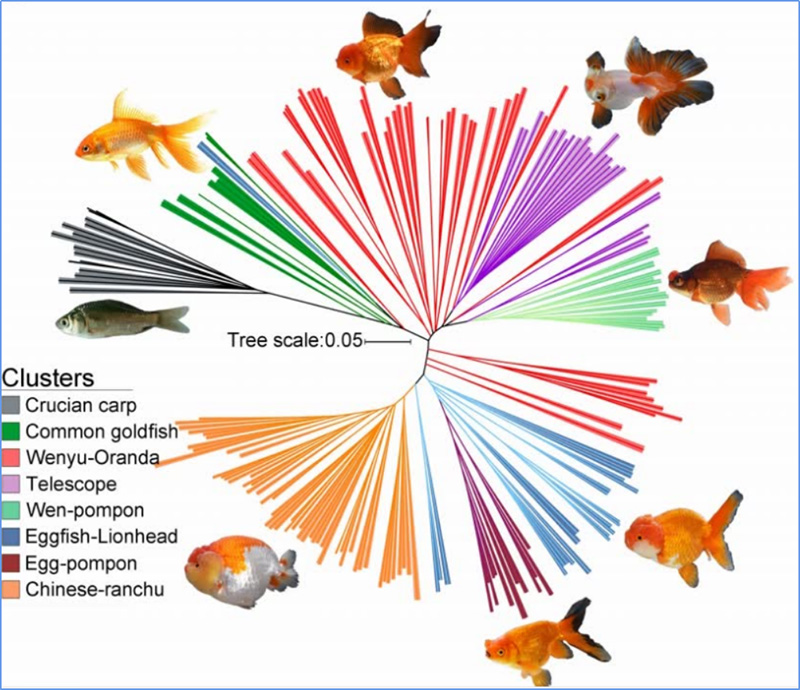
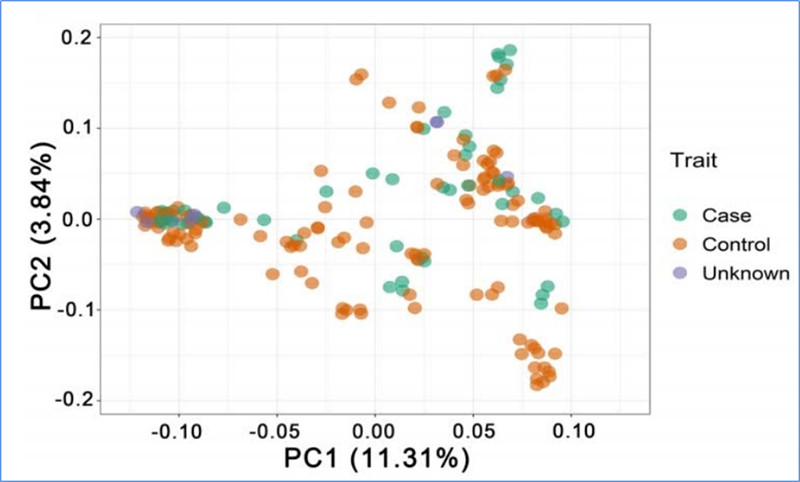
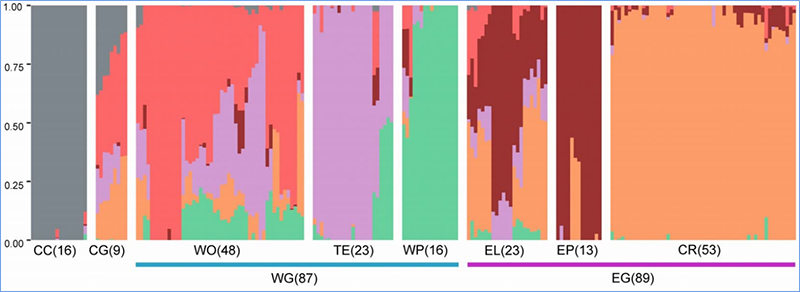
Chen, enz.al.,PNAS, 2020
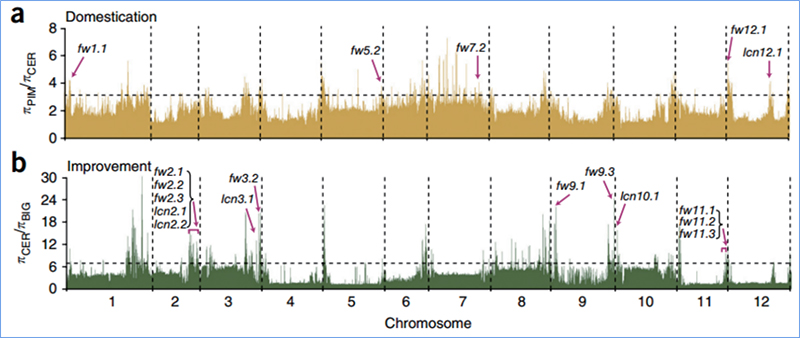
2. Selectieve sweep
Selectieve sweep verwijst naar een proces waarbij een voordelige locatie wordt geselecteerd en de frequenties van gekoppelde neutrale locaties worden verhoogd en die van niet-gekoppelde locaties worden verlaagd, wat resulteert in een vermindering van de regionale frequenties.
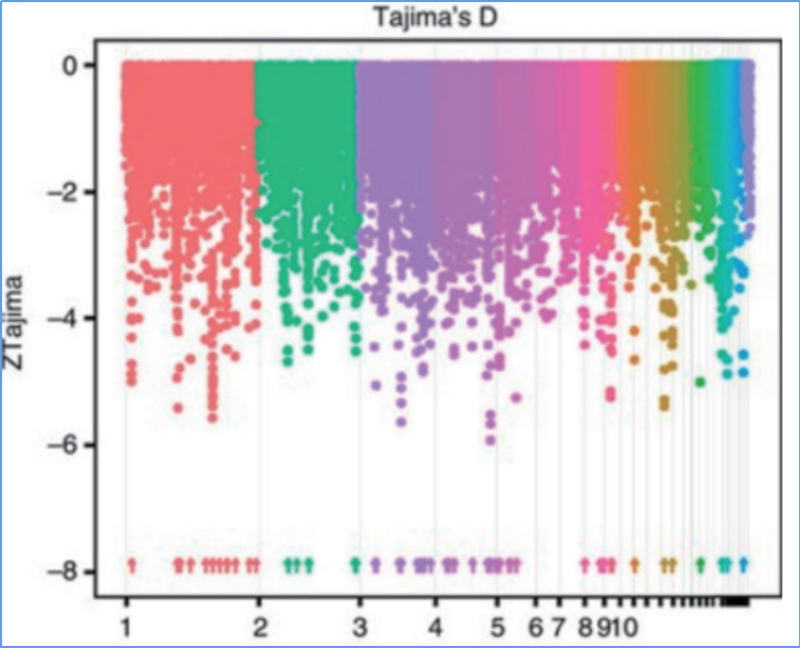
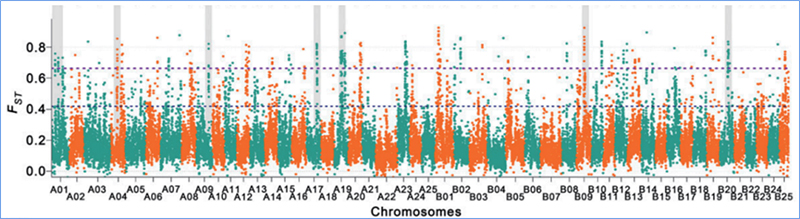
Genoombrede detectie op selectieve sweep-regio's wordt verwerkt door het berekenen van de genetische populatie-index (π, Fst, Tajima's D) van alle SNP's binnen een glijdend venster (100 Kb) bij een bepaalde stap (10 Kb).
Nucleotidendiversiteit (π)
Tajima's D
Fixatie-index (Fst)
Wu, enz.al.,Moleculaire plant, 2018
3. Genenstroom
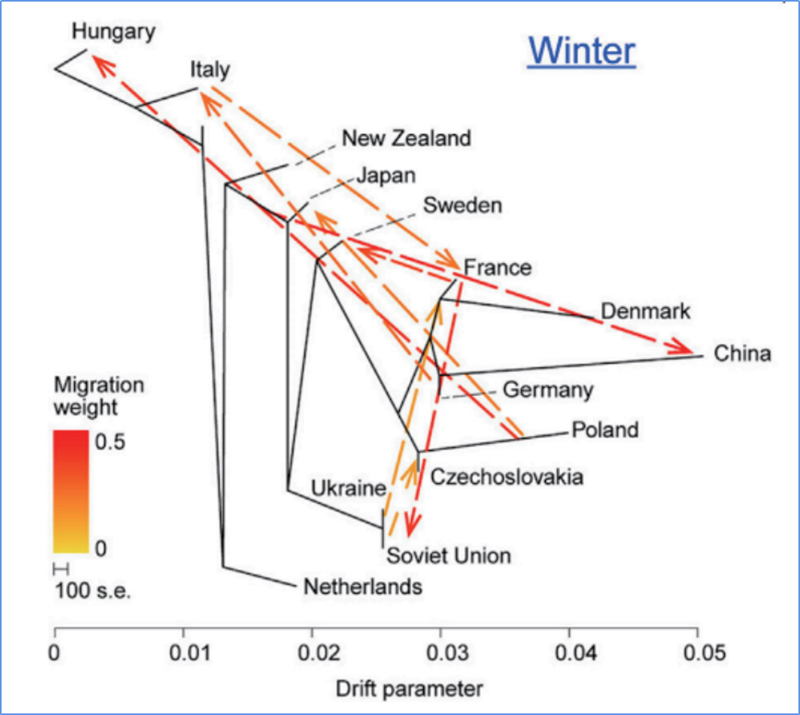
Wu, enz.al.,Moleculaire plant, 2018
4.Demografische geschiedenis
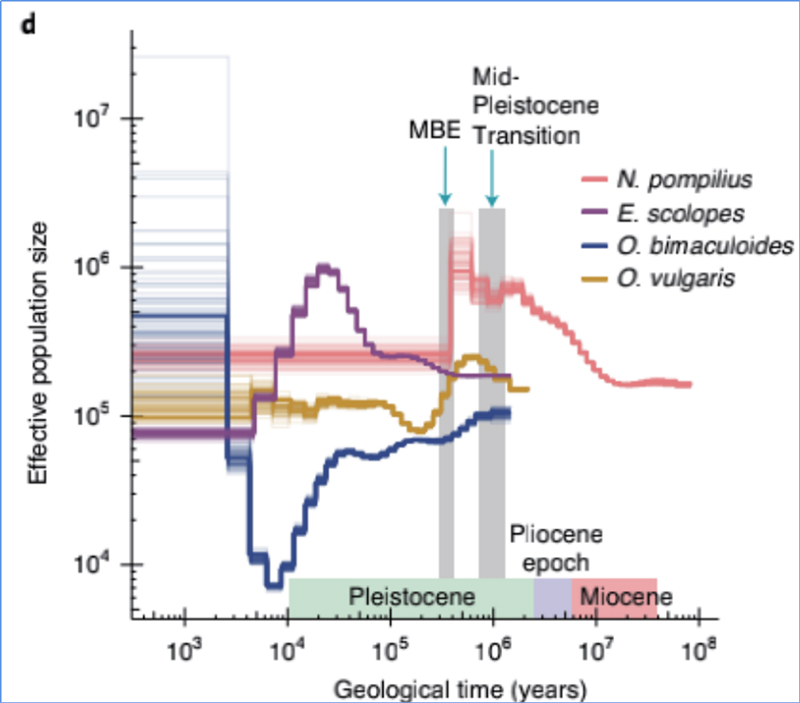
Zhang, enz.al.,Natuurecologie en evolutie, 2021
5. Divergentietijd
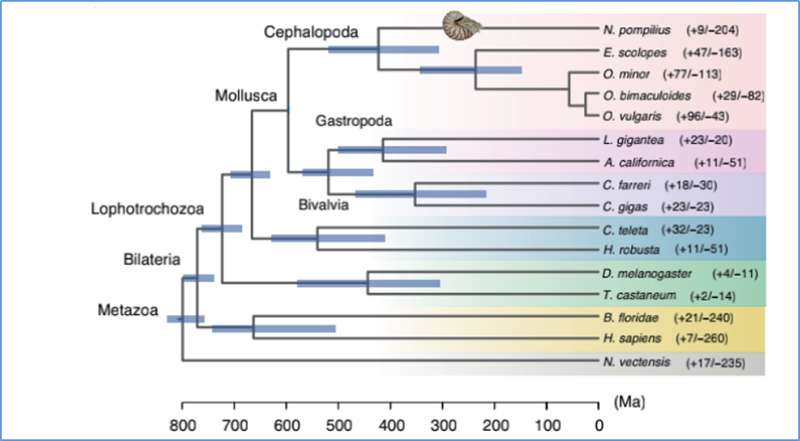
Zhang, enz.al.,Natuurecologie en evolutie, 2021
BMK-zaak
Een genomische variatiekaart biedt inzicht in de genetische basis van de selectie van Chinese lentekool (Brassica rapa ssp. Pekinensis)
Gepubliceerd: Moleculaire plant, 2018
Sequentiestrategie:
Herschikking: sequentiediepte: 10×
Belangrijkste resultaten
In dit onderzoek werden 194 Chinese kool verwerkt voor hersequencing met een gemiddelde diepte van 10×, wat 1.208.499 SNP’s en 416.070 InDels opleverde.Fylogenetische analyse van deze 194 lijnen toonde aan dat deze lijnen kunnen worden onderverdeeld in drie ecotypen: lente, zomer en herfst.Bovendien gaven de populatiestructuur en PCA-analyse aan dat Chinese lentekool afkomstig was van een herfstkool in Shandong, China.Deze werden vervolgens geïntroduceerd in Korea en Japan, gekruist met lokale lijnen en enkele laatgroeiende variëteiten ervan werden terug geïntroduceerd in China en werden uiteindelijk Chinese lentekool.
Genoombrede scanning van Chinese lentekolen en herfstkolen bij selectie onthulde 23 genomische loci die een sterke selectie hebben ondergaan, waarvan er twee overlapten met de controleregio voor de vluchttijd op basis van QTL-mapping.Deze twee regio's bleken sleutelgenen te bevatten die de bloei reguleren, BrVIN3.1 en BrFLC1.Door transcriptoomonderzoek en transgene experimenten werd verder bevestigd dat deze twee genen betrokken zijn bij de schiettijd.
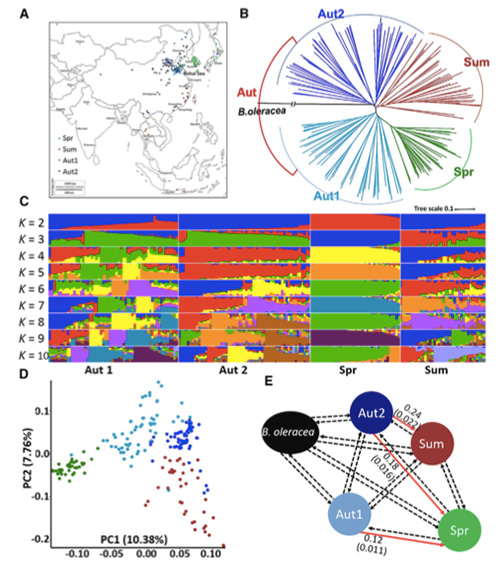 Populatiestructuuranalyse van Chinese kool | 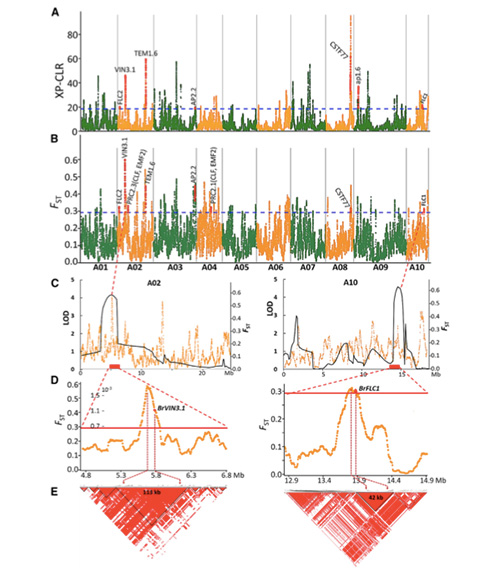 Genetische informatie over Chinese koolselectie |
Tongbing, et al."Een genomische variatiekaart biedt inzicht in de genetische basis van de selectie van Chinese lentekool (Brassica rapa ssp.pekinensis)."Moleculaire planten,11(2018):1360-1376.





