

GWAS
タイトル: 全ゲノム再配列によりセイヨウアブラナの起源とその改良に関与する遺伝子座が明らかに
ジャーナル: ネイチャーコミュニケーションズ
NGS |WGS |リシーケンス |GWAS |トランスクリプトーム |RNA配列 |アブラナ科ナパス進化 |家畜化
この研究では、Biomarker Technologies は NGS シーケンシングに関するサービスと、シーケンシング データのバイオインフォマティクス分析に関する技術サポートを提供しました。
背景
アブラナ属(菜種)は重要な油糧種子作物であり、倍数体の種分化、進化、選択のプロセスを研究するための優れたモデルです。しかし、野生種または家畜化されたドナーが親の祖先であったかどうか、および菜種の栽培化と改良に寄与した遺伝子はまだ不明です。
材料と方法
材料:588B.ナパスこの研究にはアジアから 466 件、ヨーロッパから 102 件、北米から 13 件、オーストラリアから 7 件の登録が含まれていました。成長習慣の記録に基づいて、これらの材料は 3 つのエコタイプに分類されました。春(86 アクセッション)、冬(74 アクセッション)、半冬(428 アクセッション)。
シーケンス:平均すると約5倍(3.37倍から7.71倍の範囲)
シーケンスプラットフォーム:イルミナ ハイセック 4000
データ制作:4.03 Tb のクリーン データ
SNP コール:BWA + GATK。5,294,158 個の SNP と 1,307,151 個の InDel が得られました。
結果
B.ナパスの起源
B.ナパスヨーロッパのカブの祖先から進化したサブゲノム。ヨーロッパのカブからカブへの遺伝子流動イベントB.ナプ■ サブゲノムは約 106 ~ 1170 年前に発生しました。B.ナパスC サブゲノムは、これらの系統の共通の祖先から進化した可能性があります。の祖先B.ナパスB. oleracea の 4 つの亜種の共通の祖先から分かれ、最近の遺伝子流入によりB.ナパス~108~898年前。B.ナパスC サブゲノムは、A サブゲノムよりも複雑な起源を持っています。2 つのサブゲノムの両方で強力なボトルネックが発生しました。B.ナパス進化。冬と半冬B.ナパス生態型は約 60 年前に分岐しましたが、冬と春はB.ナパス約416年前に分岐し、油糧種子と非油糧種子B.ナパス約277年前に分岐しました。
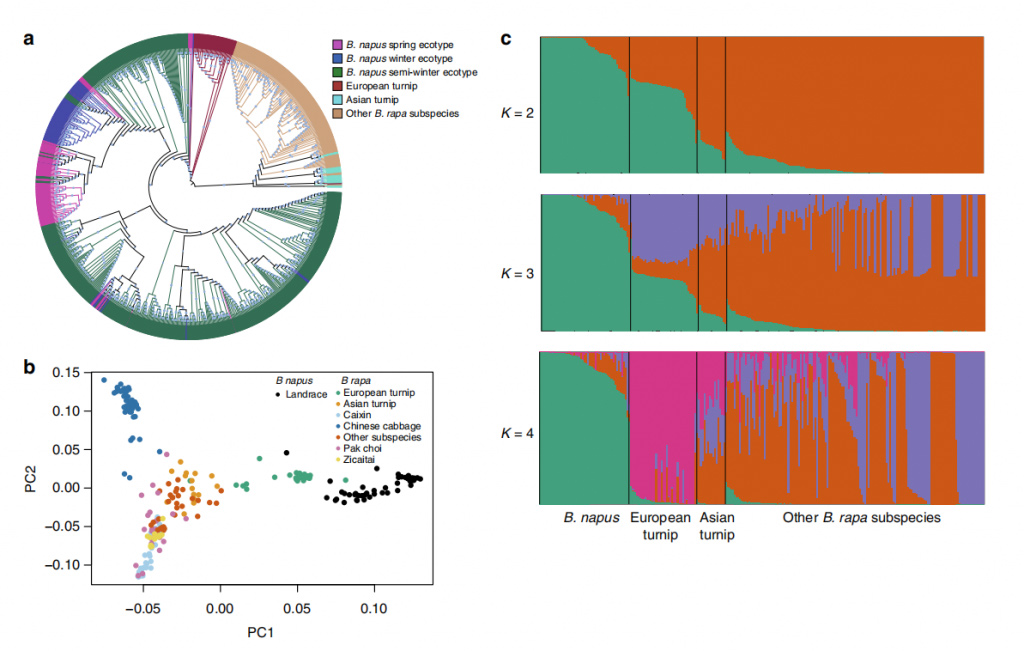
図 2 B. napus の登録数 588 件と B. rapa の登録数 199 件の集団構造。
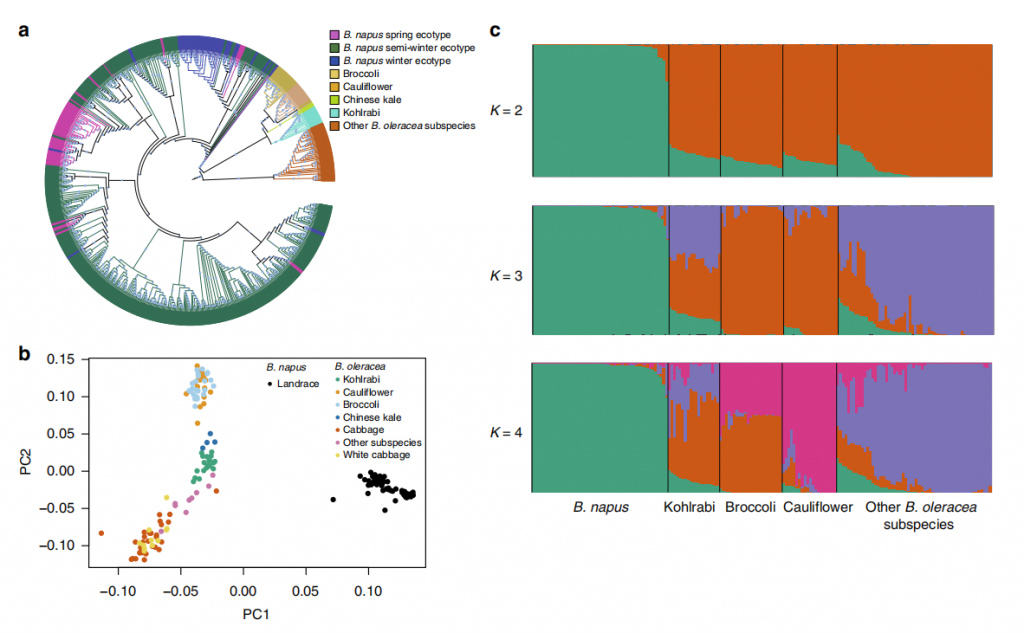
図 3 B. napus 登録 588 件と B. oleracea 登録 119 件の集団構造
選択シグナルとゲノム全体の関連研究。
改善の第 1 段階 (FSI) では、B. napus C サブゲノムでは A サブゲノムよりも多くの遺伝的多様性が失われました。FSI では、改善の第 2 段階 (SSI) よりも遺伝的分化が起こりませんでした。SSI 選択シグナル領域の遺伝子には、ストレス耐性、発達、代謝経路が豊富に含まれていました。10 の標的形質と有意に関連する 60 の遺伝子座が同定され、そのうち 5 遺伝子座は種子収量、3 遺伝子座は長角長角、3 遺伝子座は油含量、48 遺伝子座は種子の品質に関連していました。
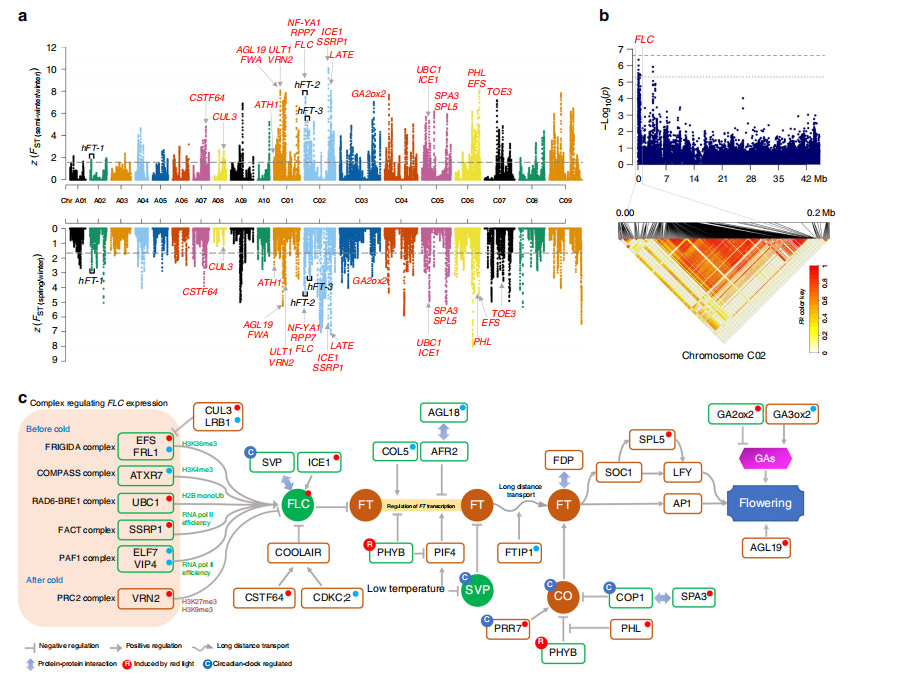
図4 セイヨウナパスのSSI中の選択された領域のゲノムワイドスキャンとアノテーション
トランスクリプトーム解析
高油分でダブルローの品種と、低油分でダブルハイの品種の 11 組織の RNAseq データにより、グルコシノレート生合成プロセスに関連する遺伝子が有意に過剰に存在することが特定されました。
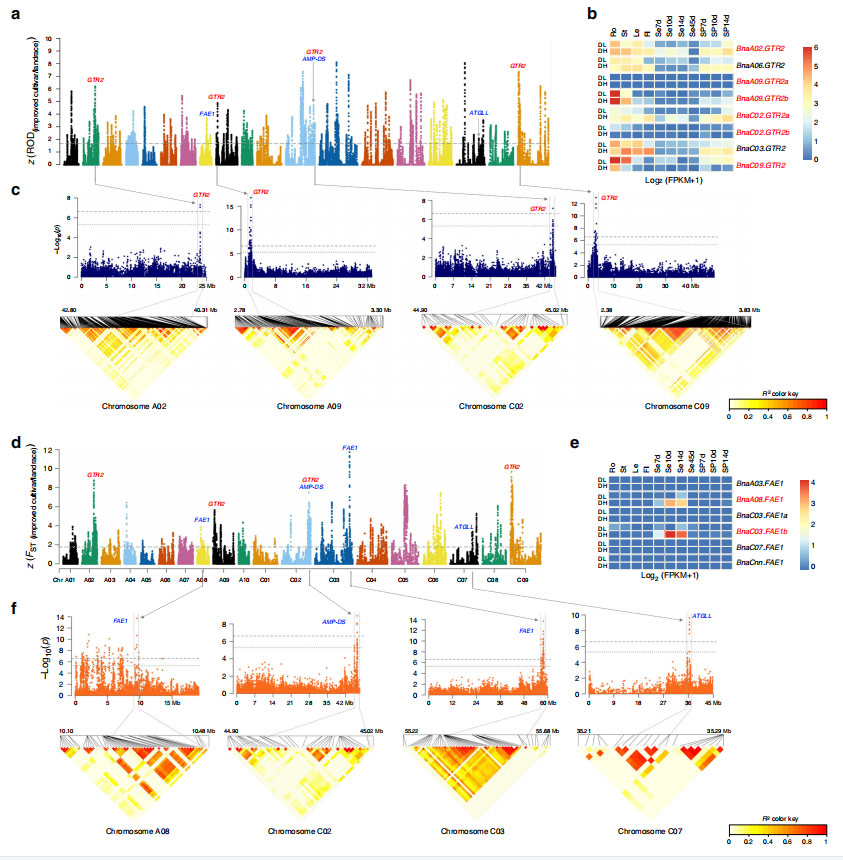
図5 セイヨウナパスの生態型改良選定による開花時期規制の概要
議論
この研究は、その起源と改善の歴史を理解するための貴重なリソースを提供しました。B.ナパスそして、重要な農学的複合形質の遺伝的基盤の分析が容易になります。有利な変異体、選択シグナル、および候補遺伝子に関連する重要な SNP は、将来、特にこの最近の異質倍数体作物とその近縁種の収量、種子の品質、油含有量、および適応性の向上に大きく貢献すると考えられます。
参照
全ゲノムの再配列により、セイヨウアブラナの起源とその改良に関与する遺伝子座が明らかになる[J]。ネイチャーコミュニケーションズ、2019年10(1)。
ニュースとハイライト この研究は、最新の成功事例をバイオマーカー テクノロジーズと共有し、新たな科学的成果や研究中に適用された著名な技術を把握することを目的としています。
投稿時刻: 2022 年 1 月 5 日