

Evolúciós genetika









Szolgáltatás előnyei
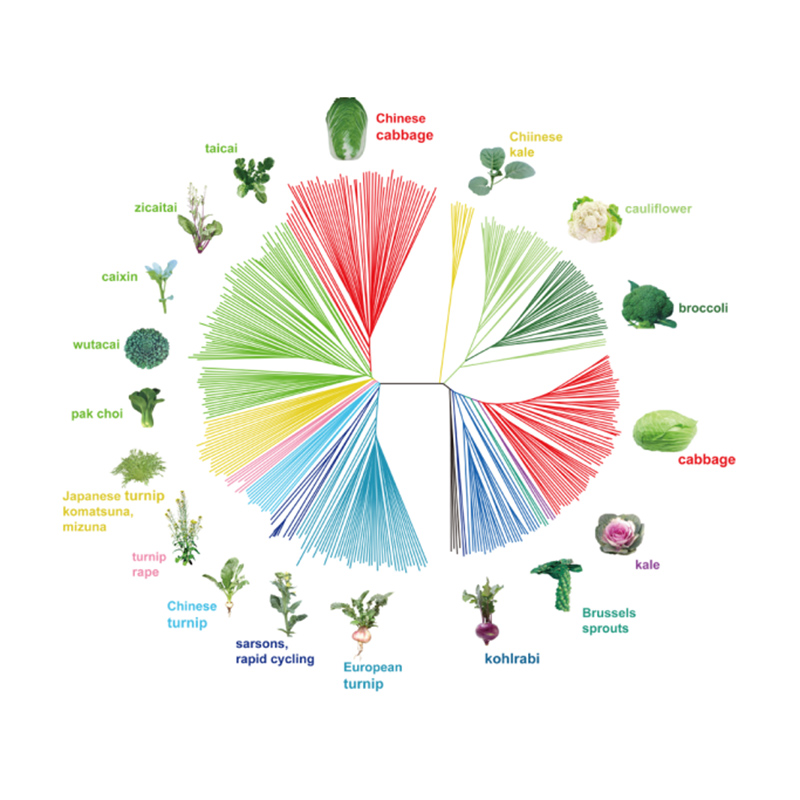
Takagi és társai,A növénynapló, 2013
● A fajok eltérési idejének és sebességének becslése nukleotid- és aminosavszintű eltérések alapján
● Megbízhatóbb filogenetikai kapcsolat feltárása a fajok között a konvergens evolúció és a párhuzamos evolúció minimális befolyásával
● Kapcsolatok létrehozása a genetikai változások és a fenotípusok között a tulajdonságokhoz kapcsolódó gének feltárása érdekében
● A genetikai sokféleség becslése, amely tükrözi a fajok evolúciós potenciálját
● Gyorsabb átfutási idő
● Széleskörű tapasztalat: A BMK több mint 12 éve hatalmas tapasztalattal rendelkezik a populációval és az evolúcióval kapcsolatos projektekben, amelyek több száz fajra vonatkoznak stb., és több mint 80 magas szintű projektben vett részt a Nature Communications, Molecular Plants, Plant Biotechnology Journal stb.
Szolgáltatási specifikációk
Anyagok:
Általában legalább három alpopuláció (pl. alfaj vagy törzs) javasolt.Minden alpopulációnak legalább 10 egyedből kell állnia (a növények >15, ritka fajok esetén csökkenthető).
Szekvenálási stratégia:
* A WGS használható jó minőségű referenciagenommal rendelkező fajokhoz, míg az SLAF-Seq alkalmazható referenciagenommal vagy anélkül, vagy rossz minőségű referenciagenomhoz.
| A genom méretére vonatkozik | WGS | SLAF-címkék (×10 000) |
| ≤ 500 Mb | 10×/egyén | A WGS inkább ajánlott |
| 500 Mb - 1 Gb | 10 | |
| 1 Gb - 2 Gb | 20 | |
| ≥2 Gb | 30 |
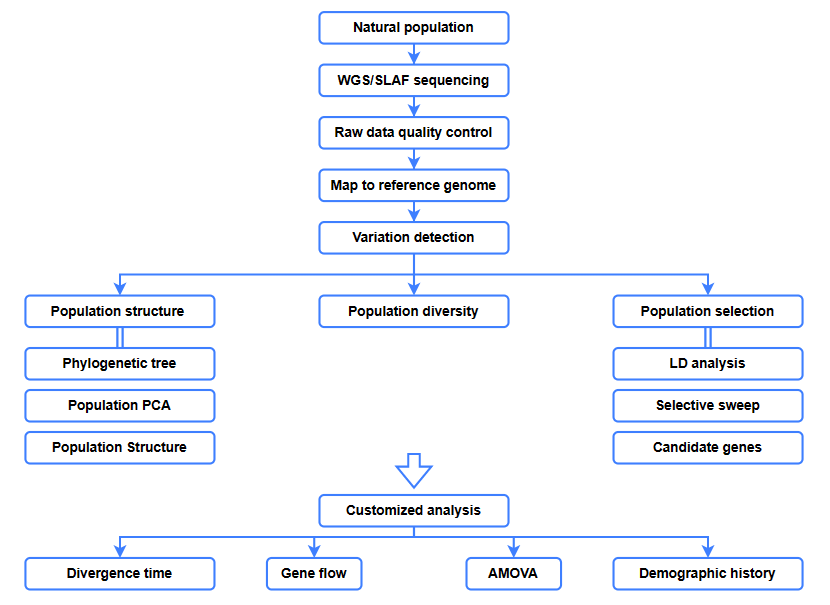
Bioinformatikai elemzések
● Evolúciós elemzés
● Szelektív söprés
● Génáramlás
● Demográfiai előzmények
● Eltérési idő
Mintakövetelmények és szállítás
Mintakövetelmények:
| Faj | Szövet | WGS-NGS | SLAF |
| Állat
| Viscerális szövet |
0,5-1 g
|
0,5 g
|
| Izomszövet | |||
| Emlős vér | 1,5 ml
| 1,5 ml
| |
| Baromfi/hal vér | |||
| Növény
| Friss Levél | 1-2 g | 0,5-1 g |
| Szirom/szár | |||
| Gyökér/Mag | |||
| Sejtek | Tenyésztett sejt |
| gDNS | Koncentráció | Összeg (ug) | OD260/OD280 |
| SLAF | ≥35 | ≥1,6 | 1,6-2,5 |
| WGS-NGS | ≥1 | ≥0,1 | - |
Szerviz munkafolyamat

Kísérleti tervezés
Mintaszállítás
Könyvtárépítés
Sorrendezés

Adatelemzés
Értékesítés utáni szolgáltatások
*Az itt látható demóeredmények mind a BMKGENE által közzétett genomokból származnak
1. Az evolúciós elemzés tartalmazza a filogenetikai fa felépítését, a populáció szerkezetét és a PCA genetikai variációk alapján történő felépítését.
A filogenetikai fa taxonómiai és evolúciós kapcsolatokat képvisel a közös ősökkel rendelkező fajok között.
A PCA célja az alpopulációk közötti közelség megjelenítése.
A populáció szerkezete genetikailag elkülönülő szubpopuláció jelenlétét mutatja az allélgyakoriság tekintetében.
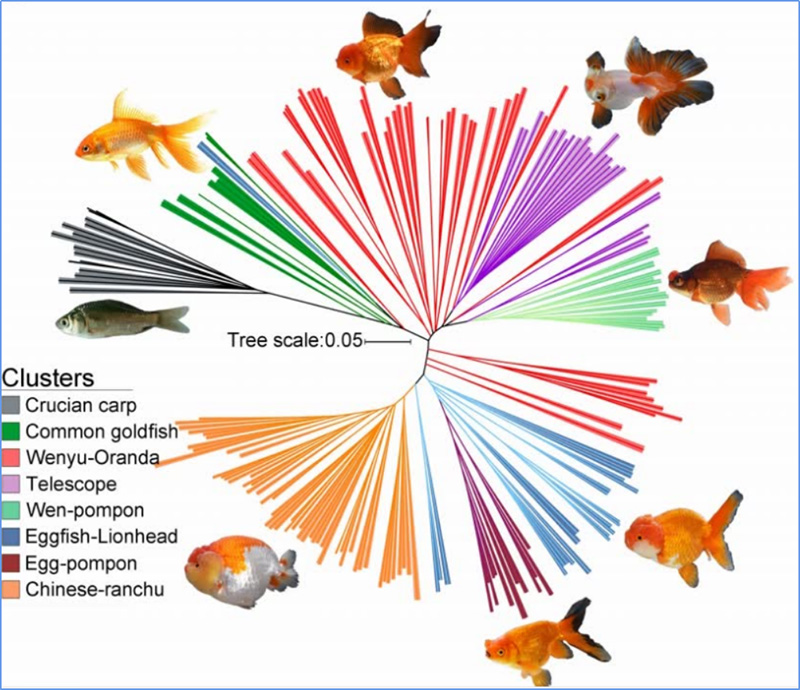
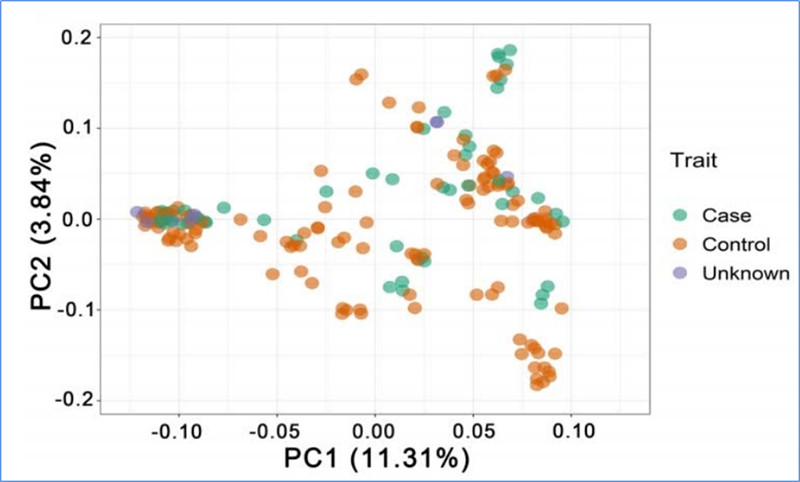
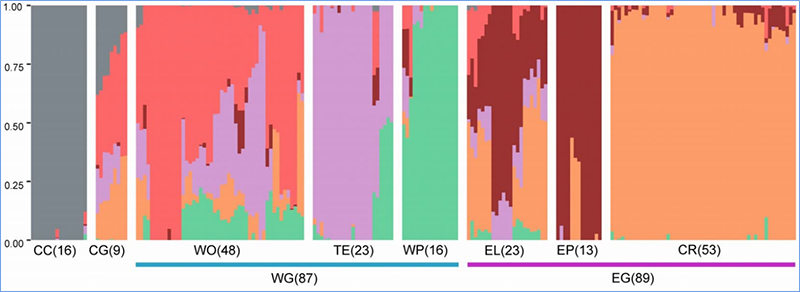
Chen és et.al.,PNAS, 2020
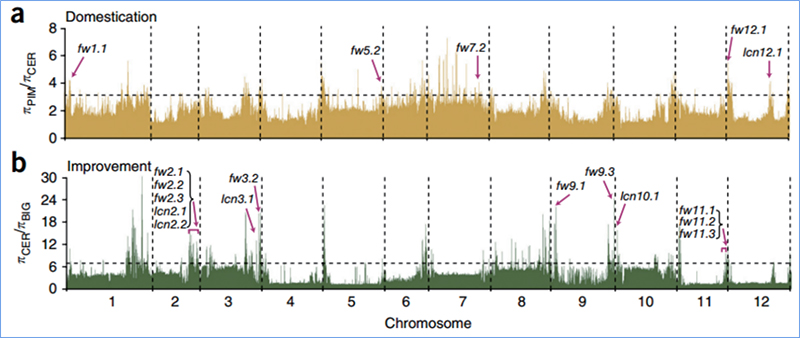
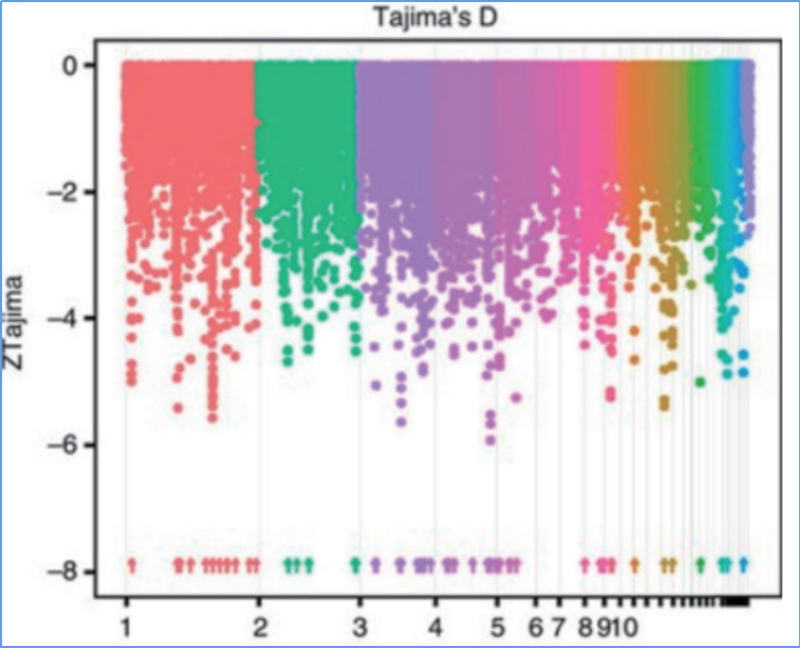
2. Szelektív seprés
A szelektív söprés egy olyan folyamat, amelynek során kiválasztanak egy előnyös helyszínt, és növelik a kapcsolt semleges helyek gyakoriságát, és csökkentik a nem kapcsolt helyek gyakoriságát, ami a regionális csökkenését eredményezi.
A szelektív sweep régiók genomszintű detektálását az összes SNP populációgenetikai indexének (π,Fst, Tajima D) kiszámításával dolgozzák fel egy csúszó ablakon (100 Kb) egy bizonyos lépésben (10 Kb).
Nukleotiddiverzitás (π)
Tajima D
Rögzítési index (Fst)
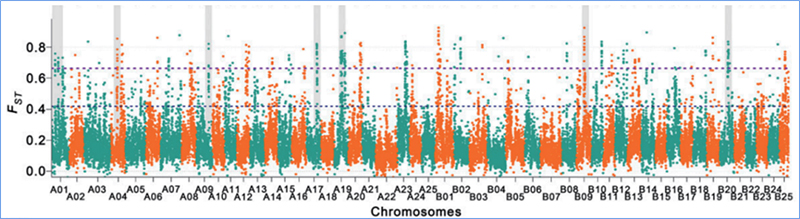
Wu és et.al.,Molekuláris növény, 2018
3.Gene Flow
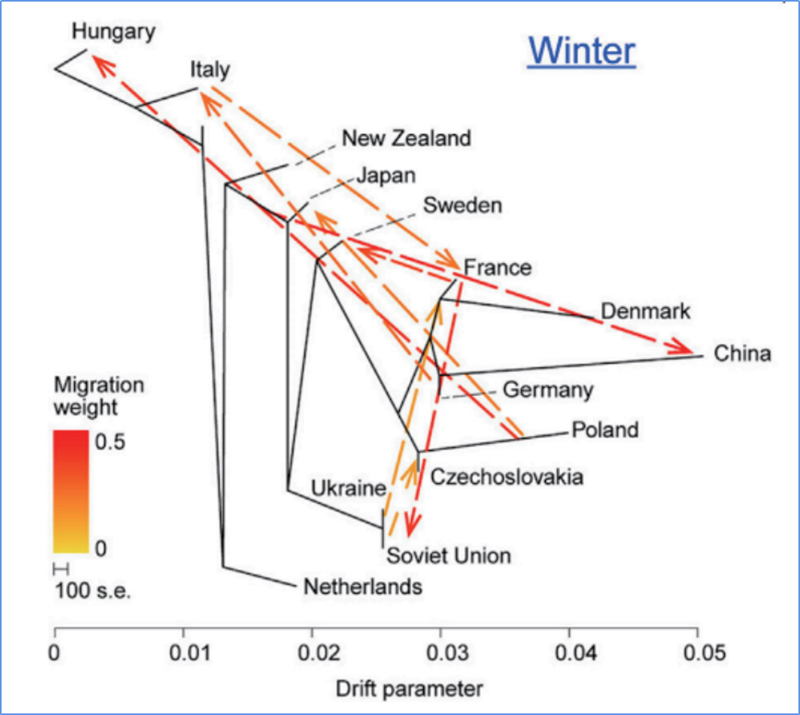
Wu és et.al.,Molekuláris növény, 2018
4. Demográfiai történelem
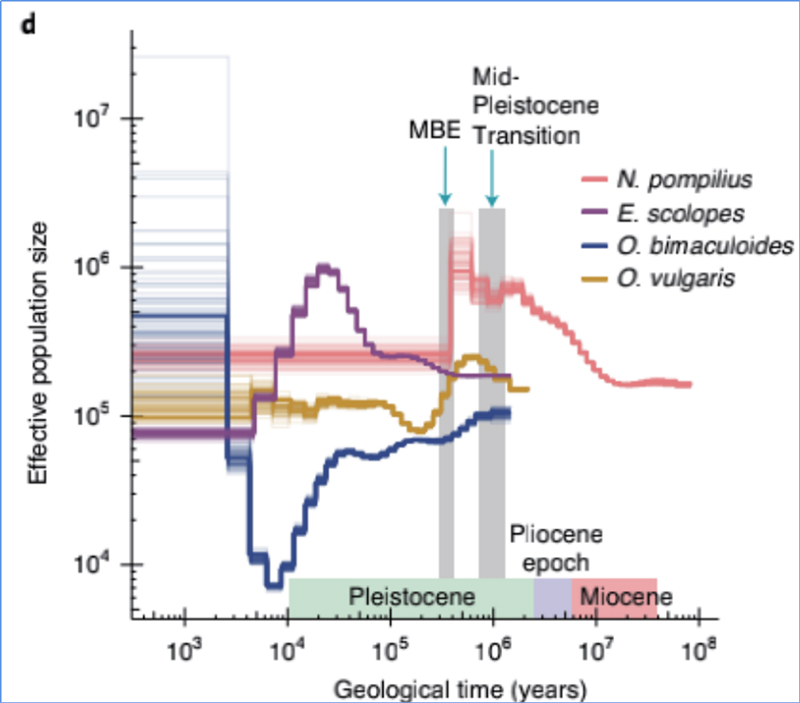
Zhang és et.al.,Természetökológia és evolúció, 2021
5. Eltérési idő
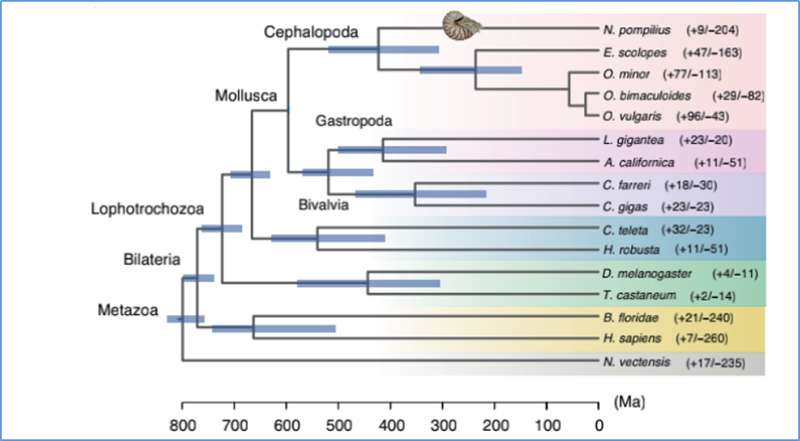
Zhang és et.al.,Természetökológia és evolúció, 2021
BMK tok
A genomi variációs térkép betekintést nyújt a tavaszi kínai káposzta (Brassica rapa ssp. Pekinensis) szelekciójának genetikai alapjaiba
Közzétett: Molekuláris növény, 2018
Szekvenálási stratégia:
Újraszekvenálás: szekvenálási mélység: 10×
Főbb eredmények
Ebben a vizsgálatban 194 kínai káposztát dolgoztak fel újraszekvenálásra, átlagosan 10-szeres mélységgel, ami 1 208 499 SNP-t és 416 070 InDelt eredményezett.A 194 vonal filogenetikai elemzése azt mutatta, hogy ezek a vonalak három ökotípusra oszthatók: tavaszi, nyári és őszi.Ezenkívül a populációszerkezet és a PCA elemzés azt mutatta, hogy a tavaszi kínai káposzta egy őszi káposztából származik Shandongban, Kínában.Ezeket később behurcolták Koreába és Japánba, keresztezték a helyi vonalakkal, és néhány késői kifejlődésű fajtájuk visszakerült Kínába, és végül tavaszi kínai káposzta lett.
A tavaszi kínai káposzta és az őszi káposzta szelekció során végzett genomszintű szkennelése 23 genomiális lókuszt mutatott ki, amelyek erős szelekción mentek keresztül, amelyek közül kettő átfedésben volt a QTL-térképezés alapján a csavarozási időt szabályozó régióval.Erről a két régióról kiderült, hogy a virágzást szabályozó kulcsgéneket, a BrVIN3.1-et és a BrFLC1-et tartalmazzák.Ezt a két gént a transzkripciós vizsgálatok és a transzgenikus kísérletek is megerősítették, hogy részt vesznek a csavarozási időben.
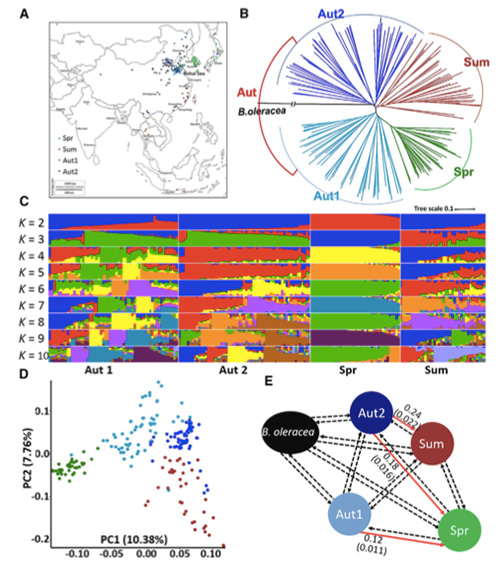 Népességszerkezeti elemzés kínai kelről | 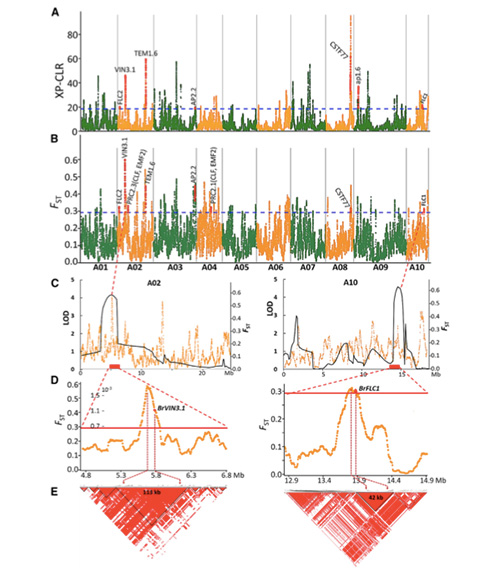 Genetikai információk a kínai kel kiválasztásáról |
Tongbing és mtsai.„A genomi variációs térkép betekintést nyújt a tavaszi kínai káposzta (Brassica rapa ssp.pekinensis) szelekciójának genetikai alapjaiba.”Molekuláris növények,11(2018): 1360-1376.





