

Evolutsiooniline geneetika









Teenuse eelised
Takagi jt,Taimede päevik, 2013
● Liikide lahknemise aja ja kiiruse hindamine nukleotiidide ja aminohapete taseme variatsioonide põhjal
● Usaldusväärsema fülogeneetilise seose paljastamine liikide vahel koos konvergentse evolutsiooni ja paralleelse evolutsiooni minimaalse mõjuga
● Seoste loomine geneetiliste muutuste ja fenotüüpide vahel, et avastada tunnustega seotud geene
● Geneetilise mitmekesisuse hindamine, mis peegeldab liikide evolutsioonipotentsiaali
● Kiirem töötlemisaeg
● Laialdased kogemused: BMK-l on üle 12 aasta kogunenud tohutu kogemus populatsiooni ja evolutsiooniga seotud projektides, hõlmates sadu liike jne ning andnud oma panuse üle 80 kõrgetasemelises projektis, mis on avaldatud ajakirjades Nature Communications, Molecular Plants, Plant Biotechnology Journal jne.
Teenuse spetsifikatsioonid
Materjalid:
Tavaliselt soovitatakse vähemalt kolme alampopulatsiooni (nt alamliiki või tüvesid).Igas alampopulatsioonis peaks olema vähemalt 10 isendit (taimed >15, võib haruldaste liikide puhul vähendada).
Järjestusstrateegia:
* WGS-i saab kasutada kvaliteetse võrdlusgenoomiga liikide puhul, samas kui SLAF-Seq on rakendatav võrdlusgenoomiga või ilma või halva kvaliteediga võrdlusgenoomiga liikide puhul.
| Kohaldatav genoomi suuruse suhtes | WGS | SLAF-sildid (×10 000) |
| ≤ 500 Mb | 10×/üksik | WGS on rohkem soovitatav |
| 500 Mb - 1 Gb | 10 | |
| 1 Gb - 2 Gb | 20 | |
| ≥2 Gb | 30 |
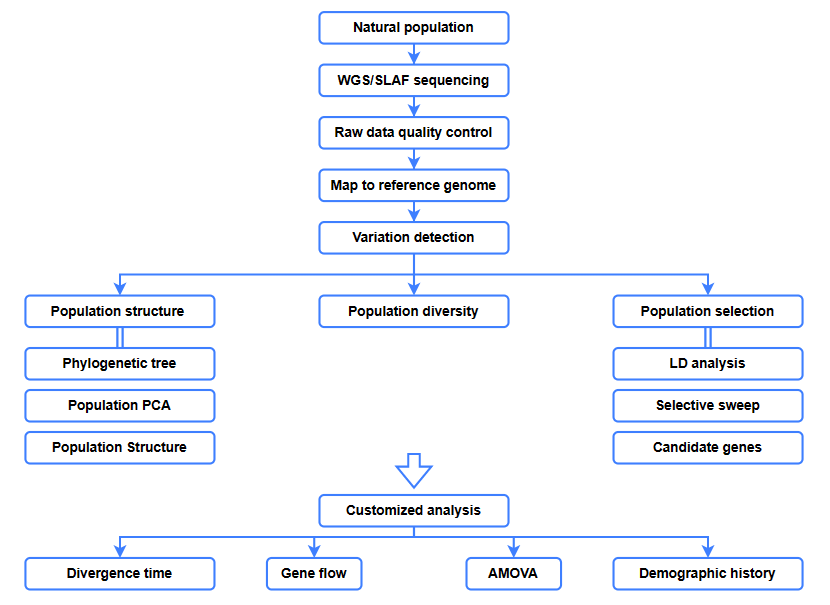
Bioinformaatika analüüsid
● Evolutsiooniline analüüs
● Valikuline pühkimine
● Geenivool
● Demograafiline ajalugu
● Divergentsi aeg
Näidisnõuded ja kohaletoimetamine
Näidisnõuded:
| Liigid | Pabertaskurätik | WGS-NGS | SLAF |
| Loom
| Vistseraalne kude |
0,5-1 g
|
0,5 g
|
| Lihaskude | |||
| Imetajate veri | 1,5 ml
| 1,5 ml
| |
| Linnuliha/kala veri | |||
| Taim
| Värske Leht | 1-2 g | 0,5-1 g |
| Kroonleht/vars | |||
| Juur/Seeme | |||
| Rakud | Kultiveeritud rakk |
| gDNA | Keskendumine | Summa (ug) | OD260/OD280 |
| SLAF | ≥35 | ≥1,6 | 1,6-2,5 |
| WGS-NGS | ≥1 | ≥0,1 | - |
Teeninduse töövoog

Katsekujundus
Näidiste kohaletoimetamine
Raamatukogu ehitus
Järjestus

Andmete analüüs
Müügijärgsed teenused
*Siin näidatud demo tulemused pärinevad kõik BMKGENE-ga avaldatud genoomidest
1.Evolutsioonianalüüs sisaldab fülogeneetilise puu, populatsiooni struktuuri ja PCA koostamist geneetiliste variatsioonide põhjal.
Fülogeneetiline puu esindab taksonoomilisi ja evolutsioonilisi suhteid ühise esivanemaga liikide vahel.
PCA eesmärk on visualiseerida alampopulatsioonide lähedust.
Populatsiooni struktuur näitab geneetiliselt eristuva alampopulatsiooni olemasolu alleelisageduste osas.
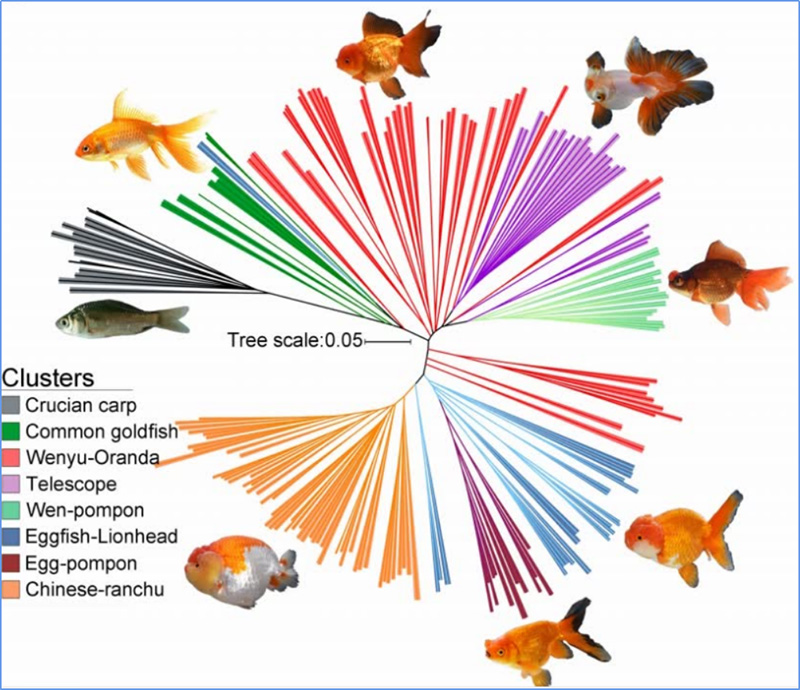
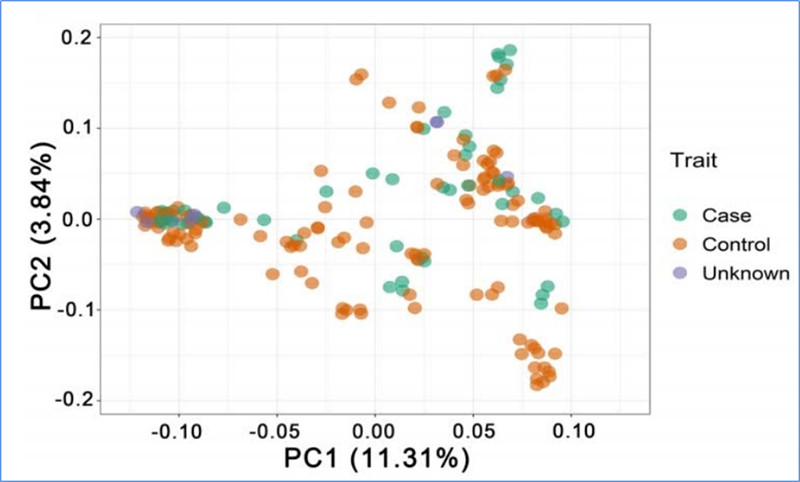
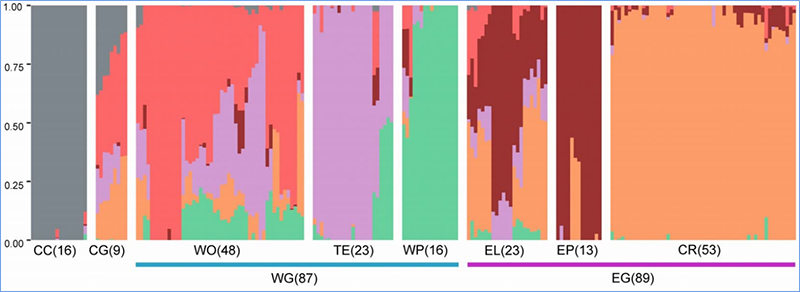
Chen et.al.,PNAS, 2020
2.Selektiivne pühkimine
Valikuline pühkimine viitab protsessile, mille käigus valitakse soodne sait ja lingitud neutraalsete saitide sagedusi suurendatakse ja sidumata saitide sagedusi vähendatakse, mille tulemuseks on piirkondliku vähenemise.
Genoomi hõlmavat tuvastamist selektiivsetes pühkimispiirkondades töödeldakse kõigi SNP-de populatsiooni geneetilise indeksi (π,Fst, Tajima D) arvutamisega libisevas aknas (100 Kb) teatud etapis (10 Kb).
Nukleotiidide mitmekesisus (π)
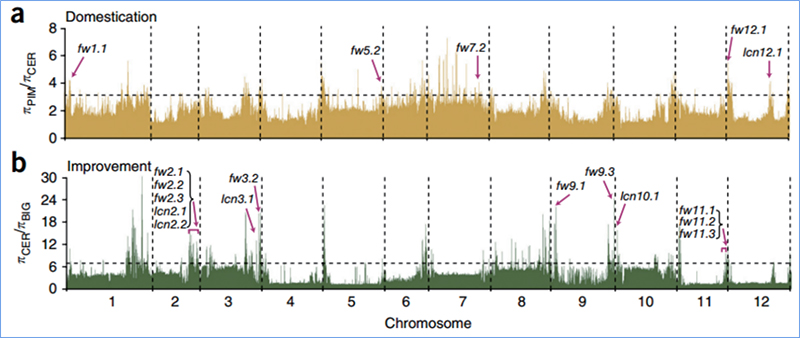
Tajima D
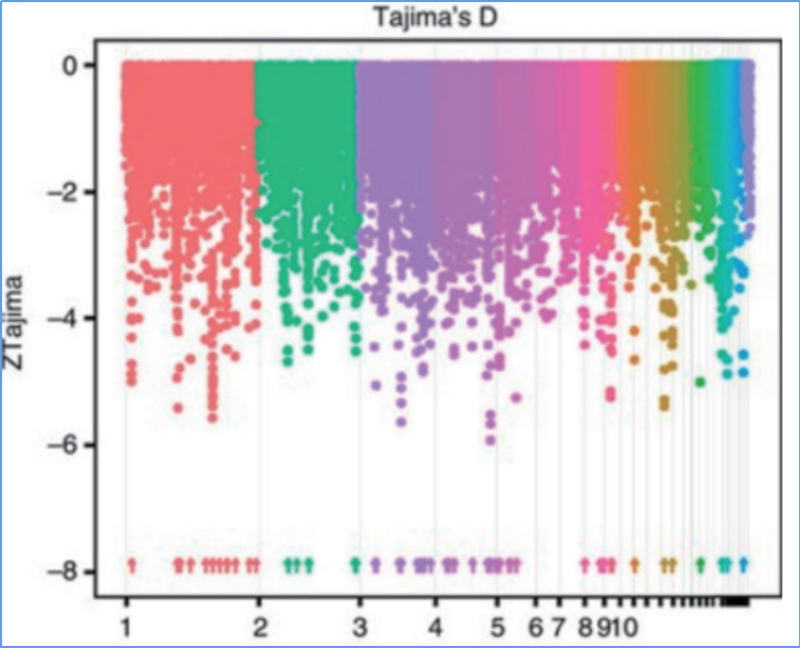
Fikseerimisindeks (Fst)
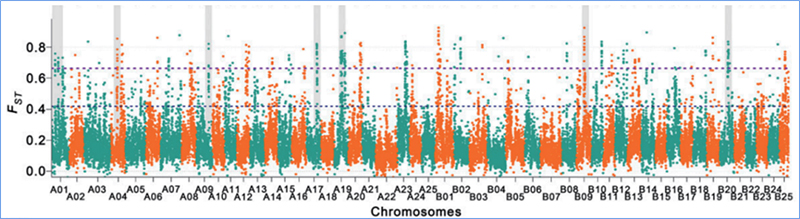
Wu et.al.,Molekulaarne taim, 2018
3.Geenivoog
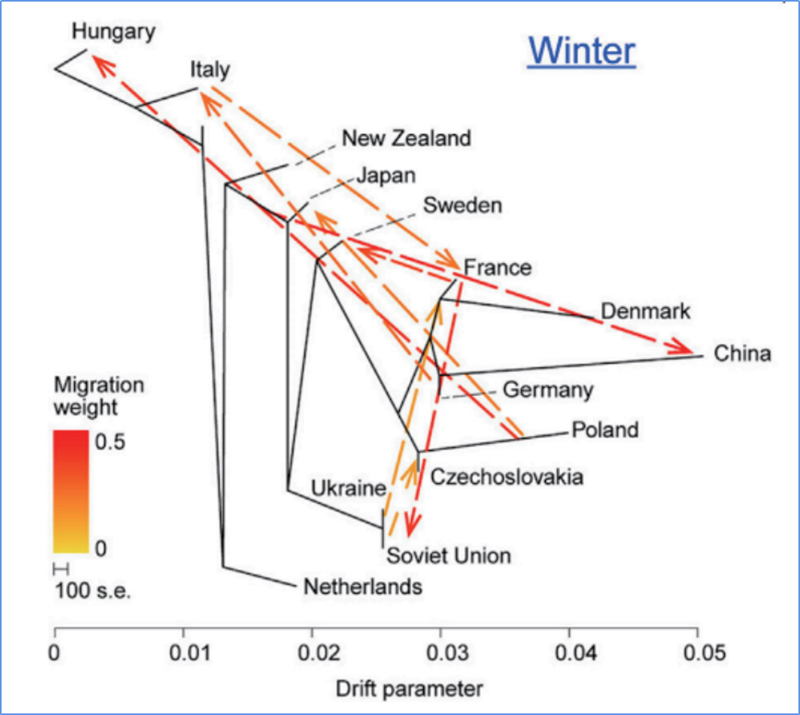
Wu et.al.,Molekulaarne taim, 2018
4.Demograafiline ajalugu
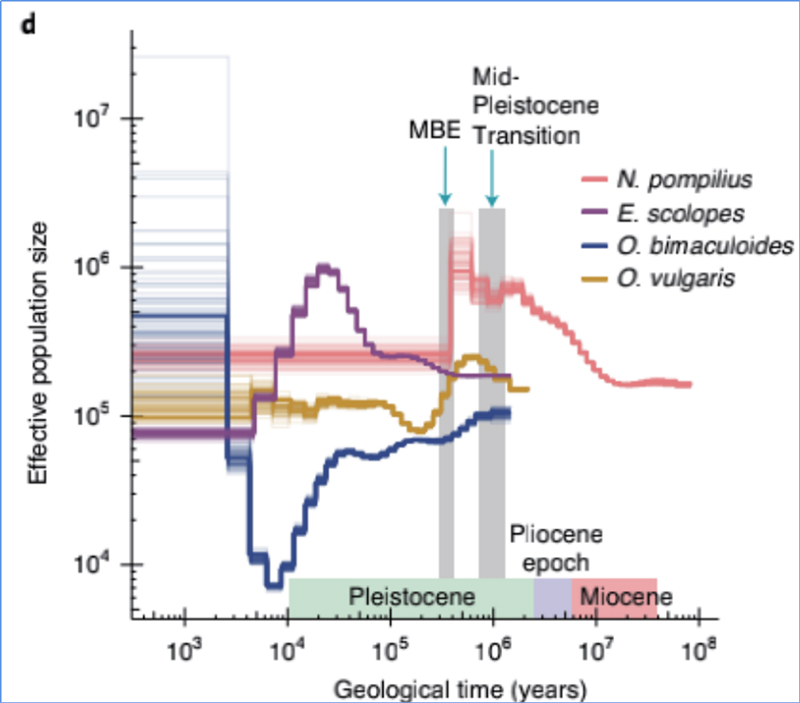
Zhang et.al.,Loodusökoloogia ja evolutsioon, 2021
5.Divergentsi aeg
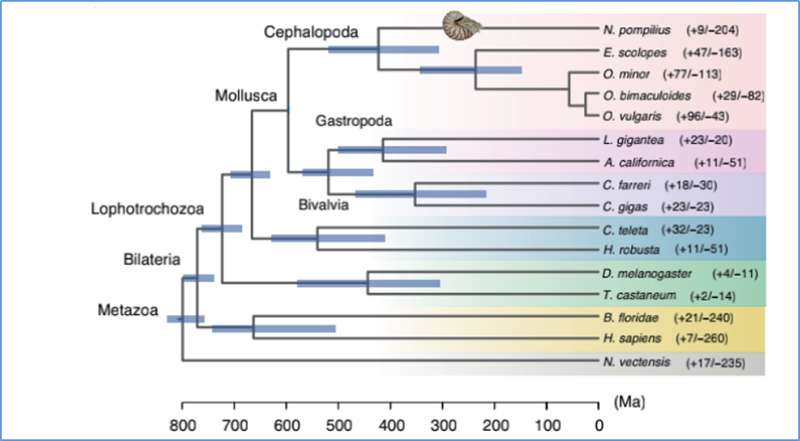
Zhang et.al.,Loodusökoloogia ja evolutsioon, 2021
BMK juhtum
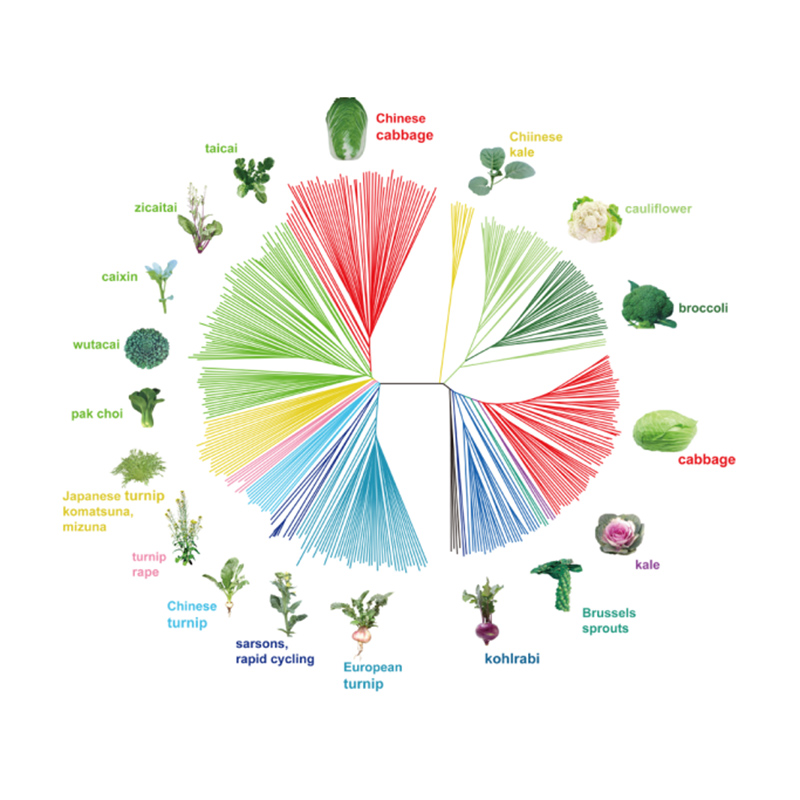
Genoomilise variatsiooni kaart annab ülevaate kevadise hiina kapsa (Brassica rapa ssp. Pekinensis) selektsiooni geneetilisest alusest.
Avaldatud: Molekulaarne taim, 2018
Järjestusstrateegia:
Kordusjärjestus: järjestuse sügavus: 10×
Peamised tulemused
Selles uuringus töödeldi 194 hiina kapsast uuesti sekveneerimiseks keskmise sügavusega 10 ×, mis andis 1 208 499 SNP-d ja 416 070 InDeli.Nende 194 liini filogeneetiline analüüs näitas, et need liinid võib jagada kolme ökotüüpi: kevad, suvi ja sügis.Lisaks näitasid populatsiooni struktuur ja PCA analüüs, et kevadine hiina kapsas pärines Hiinas Shandongis asuvast sügiskapsast.Neid toodi hiljem Koreasse ja Jaapanisse, ristati kohalike liinidega ning mõned neist viidi Hiinasse tagasi ja neist sai lõpuks kevadine hiina kapsas.
Kevadiste hiina kapsaste ja sügiskapsaste selektsioonil kogu genoomi hõlmav skaneerimine näitas 23 genoomset lookust, mis on läbinud tugeva valiku, millest kaks kattusid QTL-kaardistuse põhjal poltimisaega kontrolliva piirkonnaga.Leiti, et need kaks piirkonda sisaldavad võtmegeene, mis reguleerivad õitsemist, BrVIN3.1 ja BrFLC1.Transkriptoomiuuringud ja transgeensed katsed kinnitasid veel, et need kaks geeni on seotud polteerimisajaga.
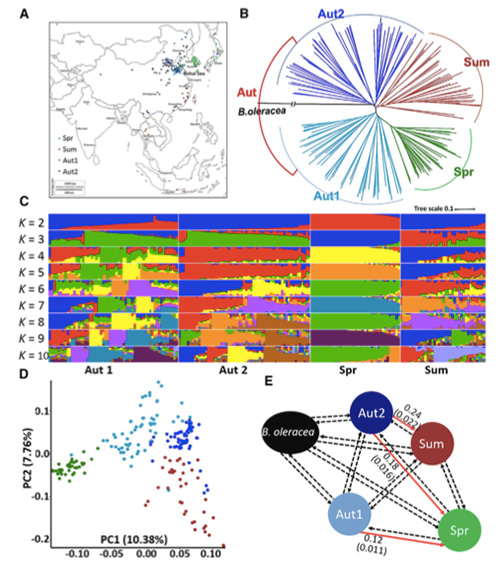 Hiina kapsa populatsioonistruktuuri analüüs | 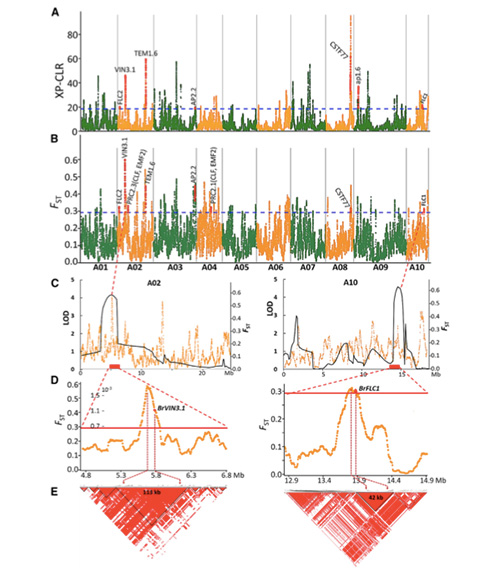 Geneetiline teave hiina kapsa valiku kohta |
Tongbing jt."Genoomiliste variatsioonide kaart annab ülevaate kevadise hiina kapsa (Brassica rapa ssp.pekinensis) valiku geneetilisest alusest."Molekulaartaimed,11 (2018): 1360-1376.





