

EVOLUCIÓN DEL GENOMA
genética de la naturaleza
Un ensamblaje genómico de alta calidad destaca las características genómicas del centeno y los genes agronómicamente importantes
PacBio |iluminar |Mapa Óptico Bionano |Ensamblaje del genoma Hi-C |Mapa genético |Barridos selectivos |Sec. de ARN |Sec. ISO |SLAF-seq
Biomarker Technologies brindó soporte técnico en la secuenciación de Pacbio, la secuenciación Hi-C y el análisis de datos en este estudio.
Reflejos
1.Se obtuvo el primer genoma de Rye de alta calidad a nivel cromosómico, que tiene un tamaño de cromosoma único superior a 1 Gb.
2. En comparación con el genoma de Tu, Aet y Hv, se observó un evento LTR-RT reciente único en el genoma de Rye, que fue responsable de la extensión del tamaño del genoma de centeno.
3. La divergencia entre el centeno y los trigos diploides tuvo lugar después de la separación de la cebada del trigo, siendo los tiempos de divergencia para los dos eventos de aproximadamente 9,6 y 15 millones de años.
La fosforilación de los genes FT puede controlar el rasgo de partida temprana en el centeno.
4. El análisis de barrido selectivo indica una posible participación de ScID1 en la regulación de la fecha de partida y su probable selección por domesticación en centeno.
Fondo
Fondo
El centeno es un valioso cultivo alimentario y forrajero, un recurso genético importante para el mejoramiento del trigo y el triticale, y un material indispensable para estudios genómicos comparativos eficientes en gramíneas.El centeno weining, una variedad de floración temprana cultivada en China, destaca por su resistencia de amplio espectro tanto al mildiú polvoriento como a la roya lineal.Para comprender las bases genéticas y moleculares de los rasgos de élite del centeno y promover los estudios genómicos y de mejoramiento en centeno y cultivos relacionados, aquí secuenciamos y analizamos el genoma del centeno Weining.
Logros
Genoma del centeno
El genoma de Rye se construyó combinando lecturas de PacBio SMRT, secuenciación de lectura corta de Illumina, así como las de captura de conformación de cromatina (Hi-C), mapeo genético y análisis BioNano.Los cóntigs ensamblados (7,74 Gb) representaron el 98,47% del tamaño estimado del genoma (7,86 Gb), con el 93,67% de los cóntigos (7,25 Gb) asignados a siete cromosomas.Los elementos repetitivos constituyeron el 90,31% del genoma ensamblado.
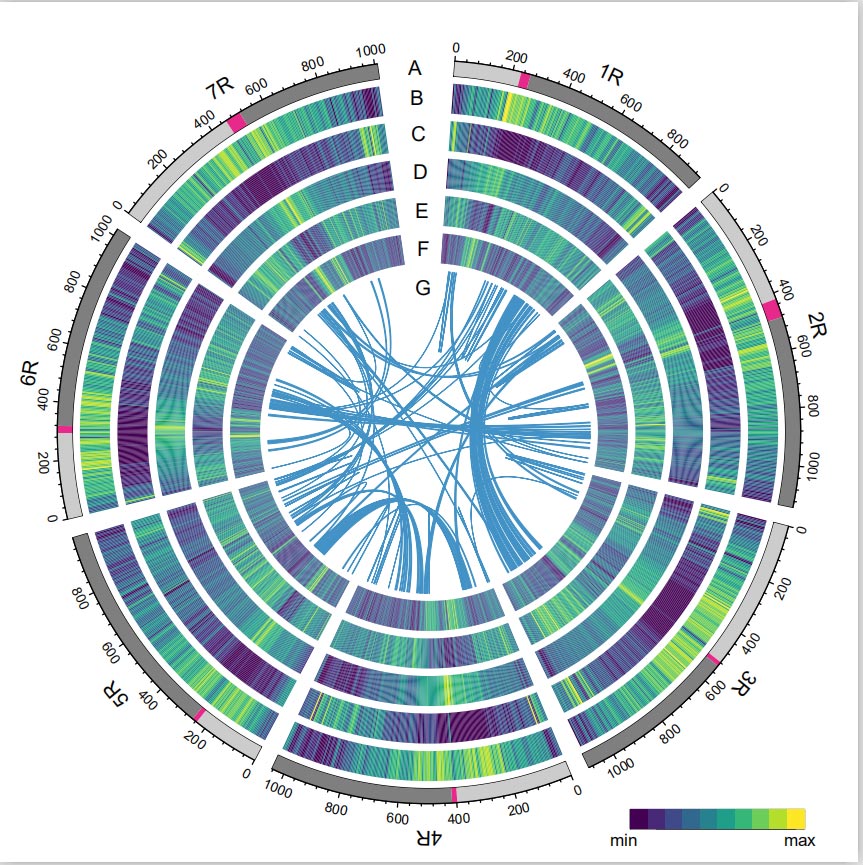
Genoma del centeno
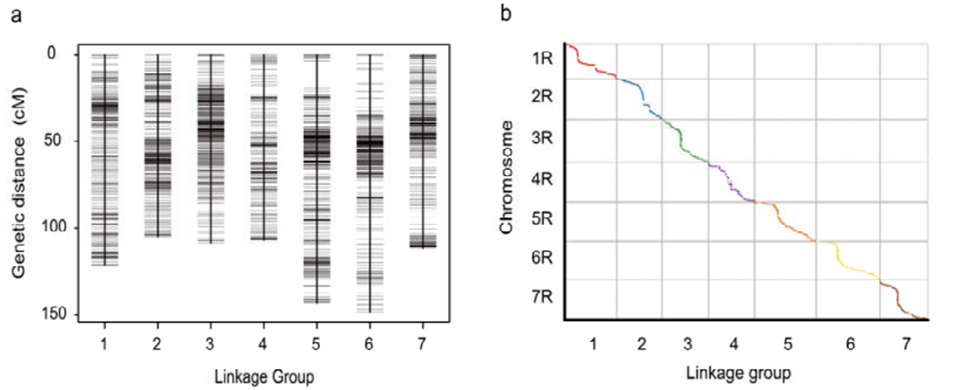
Mapa de ligamiento genético (WJ) desarrollado utilizando 295 plantas F2 derivadas del cruce de dos variedades locales de centeno (Weining × Jingzhou)
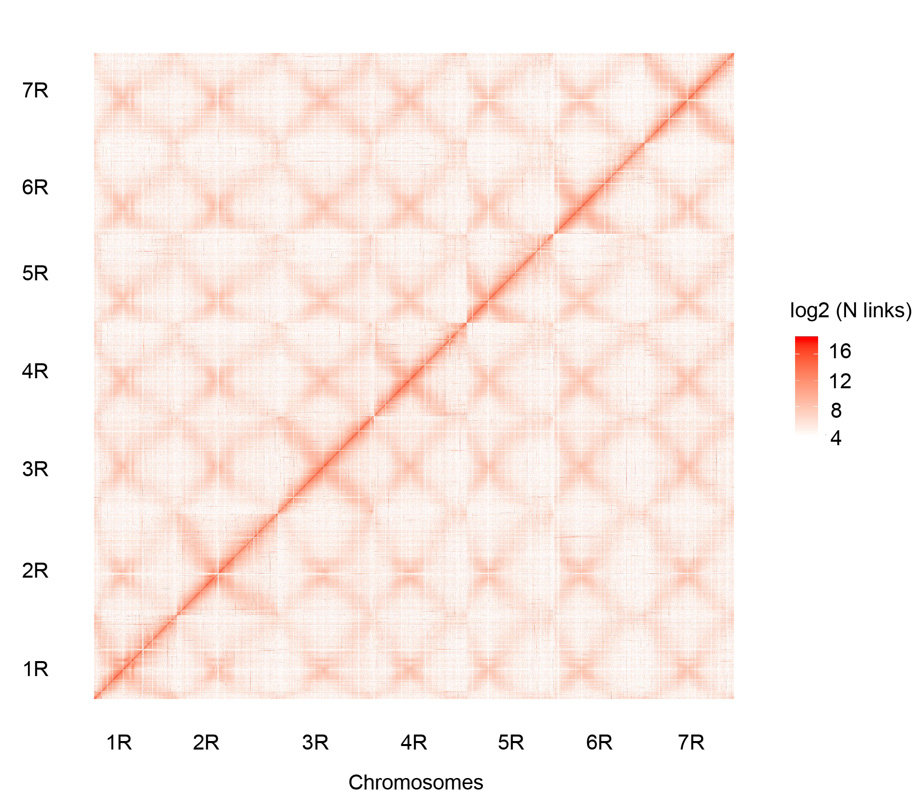
Mapa de contactos Hi-C de los siete cromosomas de centeno Weining ensamblados (1R - 7R)
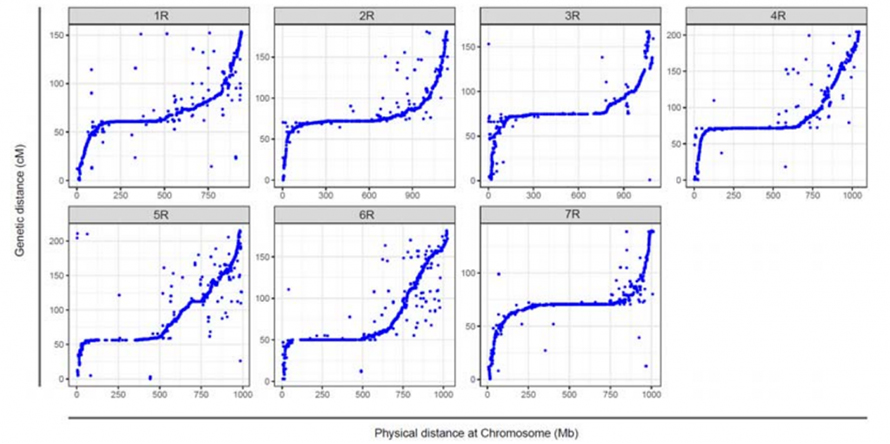
Alineación entre los siete cromosomas ensamblados del centeno Weining y los siete grupos de enlace del centeno desarrollados utilizando la población RIL Lo7 x Lo255
Se encontró que el valor del índice de ensamblaje LTR (LAI) del genoma de centeno era 18,42 y se identificaron 1.393 (96,74%) de los 1.440 genes BUSCO altamente conservados. Estos resultados sugieren que la secuencia del genoma de centeno de Weining es de alta calidad en ambos intergénicos. y regiones genéticas.Se predijo un total de 86.991 genes codificadores de proteínas, incluidos 45.596 genes de alta confianza (HC) y 41.395 genes de baja confianza (LC).
2. Análisis de TE
Análisis de TE.Un total de 6,99 Gb, que representan el 90,31% del ensamblaje de Weining, fueron anotados como TE, que incluían 2.671.941 elementos pertenecientes a 537 familias.Este contenido de TE fue claramente superior al reportado previamente para Ta (84,70%), Tu (81,42%), Aet (84,40%), WEW (82,20%) o Hv (80,80%).Los retrotransposones de repetición terminal larga (LTR-RT), incluidos Gypsy, Copia y elementos RT no clasificados, fueron los TE dominantes y ocuparon el 84,49% del contenido de TE anotado y el 76,29% del genoma de Weining ensamblado;Los transposones de ADN CACTA fueron los segundos TE más abundantes, constituyendo el 11,68% del contenido de TE anotado y el 10,55% del genoma de Weining ensamblado.
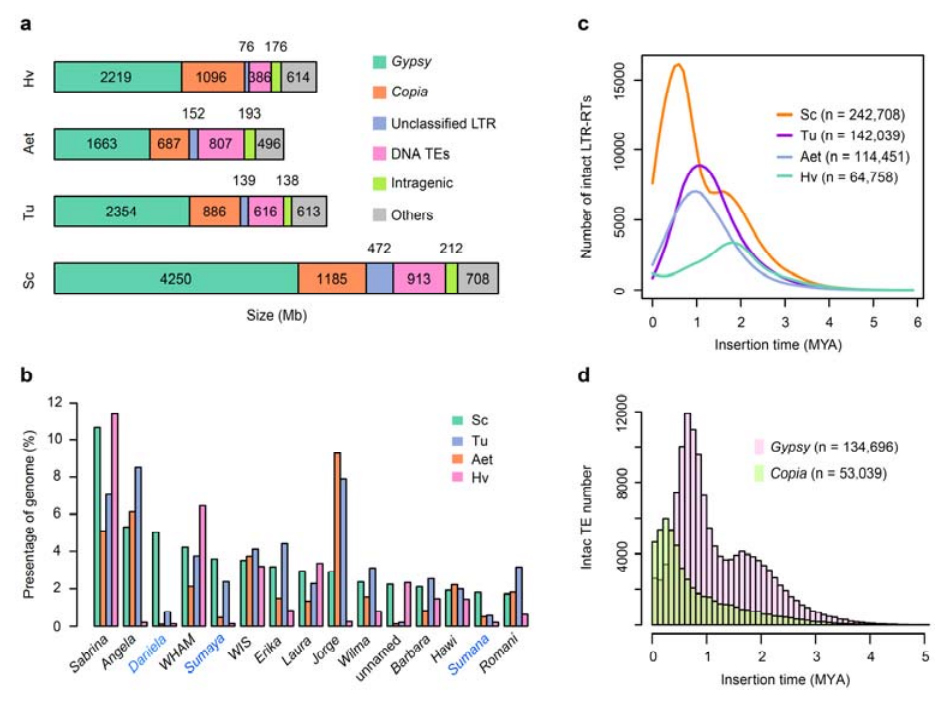
Análisis de elementos transposones de centeno.
El centeno weining tuvo una proporción comparativamente alta de inserciones recientes de LTR-RT y el pico de amplificación apareció hace alrededor de 0,5 millones de años (MYA), que fue la más reciente entre las cuatro especies;el otro pico, que se produjo hace aproximadamente 1,7 millones de años, era más antiguo y también se observó en la cebada.A nivel de superfamilia, se encontraron explosiones muy recientes de elementos Copia en centeno Weining hace 0,3 millones de años, mientras que las amplificaciones de Gypsy RT dieron forma dominante al patrón de distribución bimodal de la dinámica de explosión LTR-RT.
3. Investigación de la evolución del genoma del centeno y la sintenia cromosómica.
La divergencia entre el centeno y los trigos diploides tuvo lugar después de la separación de la cebada del trigo, siendo los tiempos de divergencia para los dos eventos de aproximadamente 9,6 y 15 millones de años, respectivamente.1R, 2R, 3R eran completamente colineales con los cromosomas de los grupos 1, 2 y 3 del trigo, respectivamente.Se encontró que 4R, 5R, 6R, 7R existen fusiones y segmentos a gran escala.
4. Análisis de duplicaciones genéticas y su impacto en los genes de biosíntesis de almidón.
En particular, el número de genes duplicados en tándem (TDG) y genes duplicados proximalmente (PDG) del centeno Weining fueron mayores que los encontrados para Tu, Aet, Hv, Bd y Os.Los genes duplicados transpuestos (TrDG) también fueron más numerosos que los encontrados específicamente para Tu y Aet.La expansión del genoma del centeno va acompañada de un mayor número de duplicaciones de genes.El aumento de las explosiones de TE en el centeno puede haber dado lugar a un número elevado de TrDG.
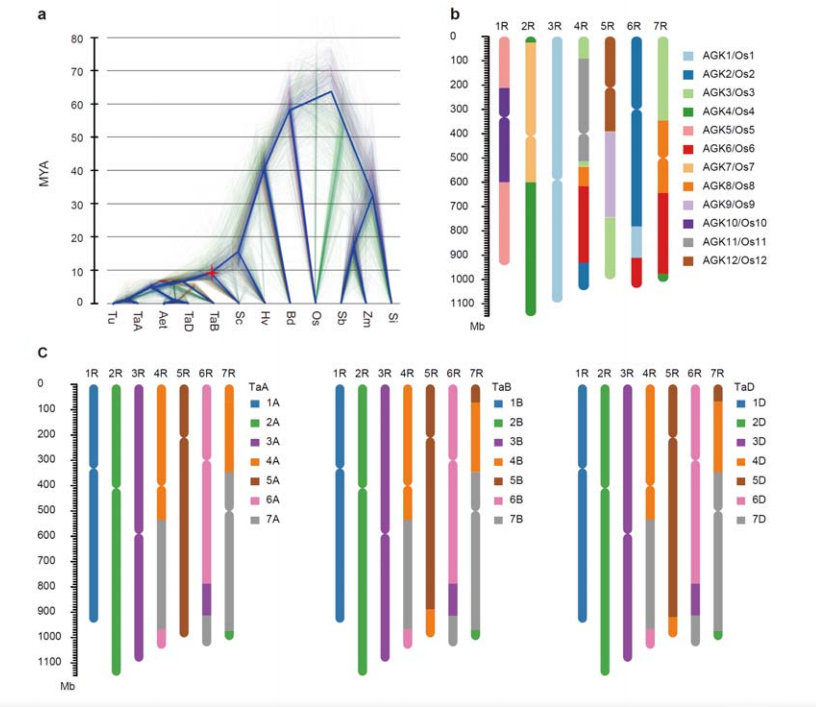
Análisis evolutivos y de sintenia cromosómica del genoma del centeno.
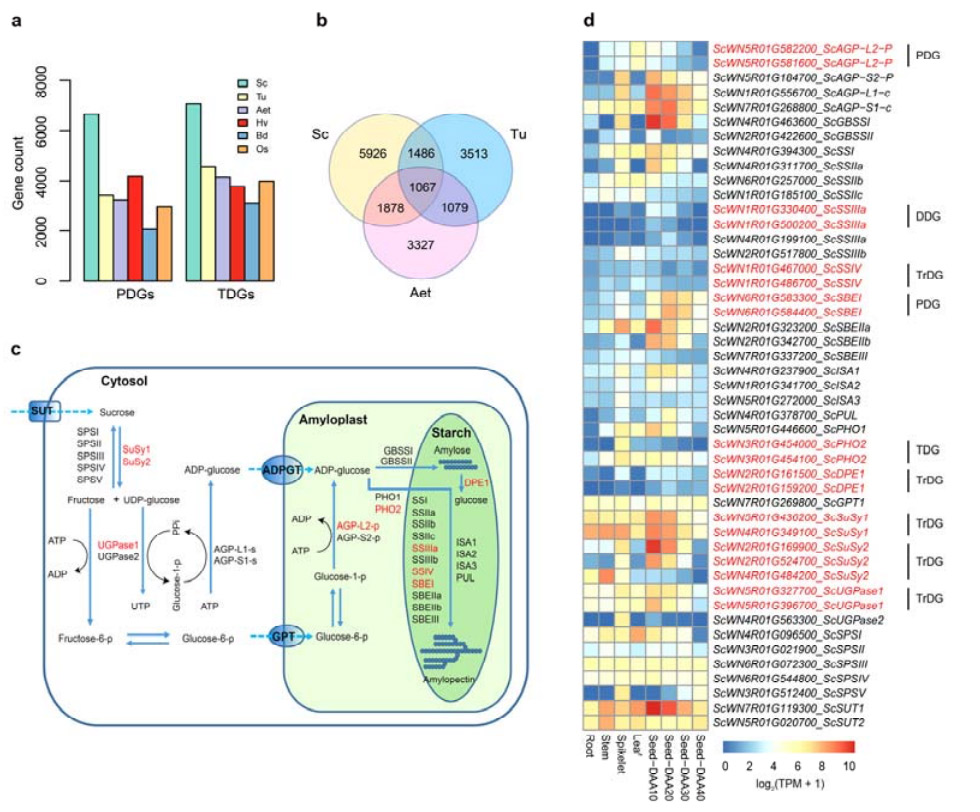
Análisis de duplicaciones de genes de centeno y su impacto en la diversidad de genes relacionados con la biosíntesis de almidón (SBRG)
5. Disección de loci del gen de la proteína de almacenamiento de semillas de centeno (SSP)
Se han identificado cuatro loci cromosómicos (Sec-1 a Sec-4) que especifican SSP de centeno en 1R o 2R.Los genes de α-gliadina evolucionaron recientemente en el trigo y en especies estrechamente relacionadas después de la divergencia del trigo del centeno.
6. Examen del factor de transcripción (TF) y genes de resistencia a enfermedades.
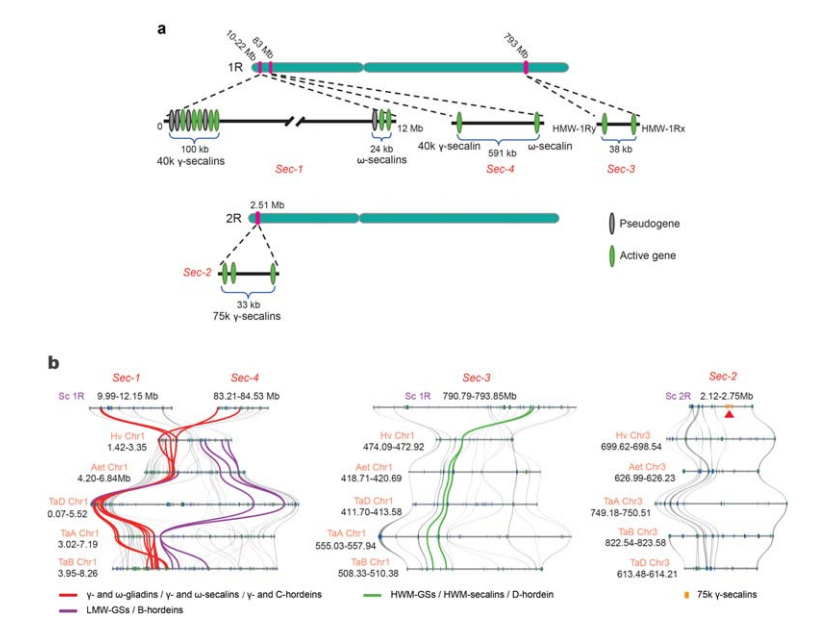
Análisis de loci de secalina de centeno.
El centeno weining tenía más genes asociados a la resistencia a enfermedades (DRA) (1989, Datos complementarios 3) que Tu (1621), Aet (1758), Hv (1508), Bd (1178), Os (1575) y A (1836). ), subgenomas B (1.728) y D (1.888) de trigo blando.
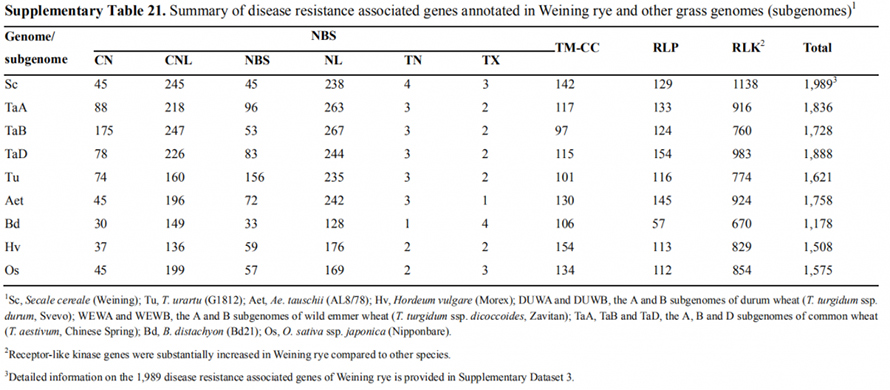
7. Investigación de las características de expresión genética asociadas con el rasgo de rumbo temprano
Se anotaron dos genes FT con expresión relativamente alta en condiciones de días largos, ScFT1 y ScFT2, en el ensamblaje del genoma de Weining.Se encontró relación con la fosforilación de dos residuos de aminoácidos de ScFT2 (S76 y T132) con la reducción del control del tiempo.
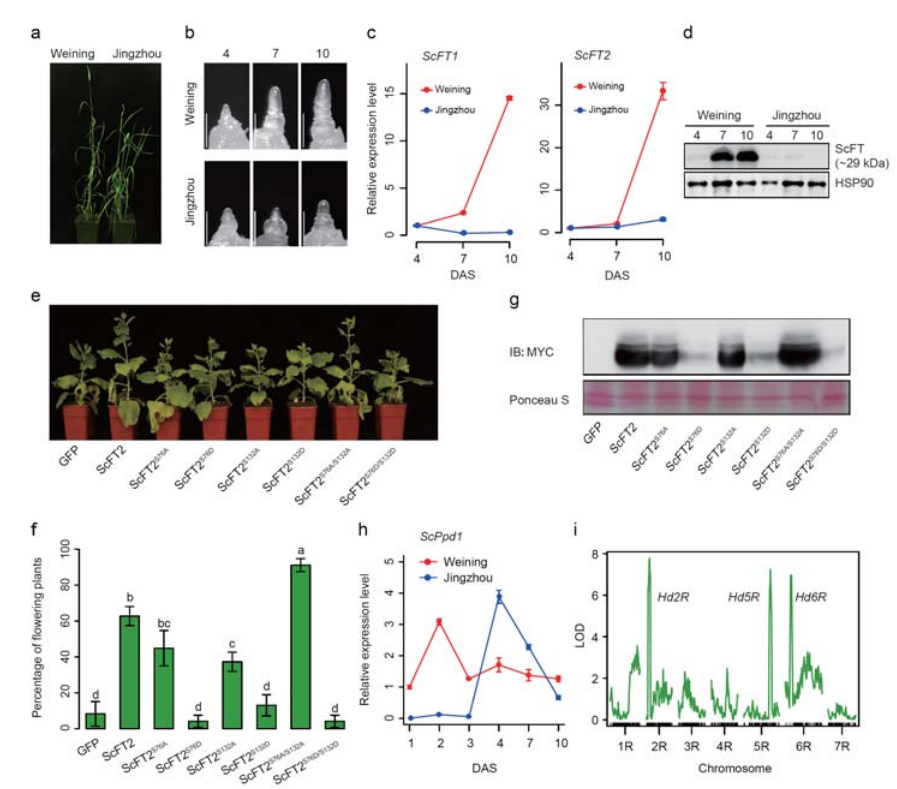
Características de desarrollo y expresión genética asociadas con el rasgo de rumbo temprano del centeno Weining
8. Minería de regiones cromosómicas y loci potencialmente involucrados en la domesticación del centeno.
Se utilizó un total de 123.647 SNP para realizar análisis de barrido selectivo entre centeno cultivado y S. vavilovii.11 señales de barrido selectivo identificadas por índice de reducción (DRI), índice de fijación (FST) y método XP-CLR.Se encontró que ScID1 podría estar involucrado en la regulación de la fecha del rumbo.
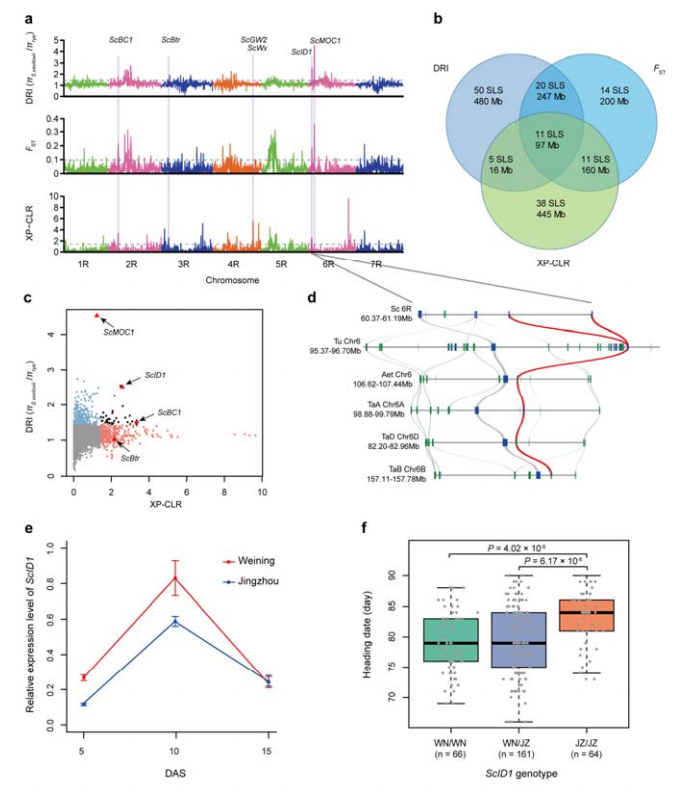
Identificación y análisis de regiones cromosómicas y loci potencialmente relacionados con la domesticación del centeno.
Referencia
Li GW et al.Un ensamblaje genómico de alta calidad resalta las características genómicas del centeno y los genes agronómicamente importantes.Genética de la naturaleza (2021)
Noticias y destacados tiene como objetivo compartir los últimos casos exitosos con Biomarker Technologies, capturando logros científicos novedosos, así como técnicas destacadas aplicadas durante el estudio.
Hora de publicación: 05-ene-2022