

Specific-Locus Amplified Fragment Sequencing (SLAF-Seq)





Servicedetaljer
Teknisk ordning
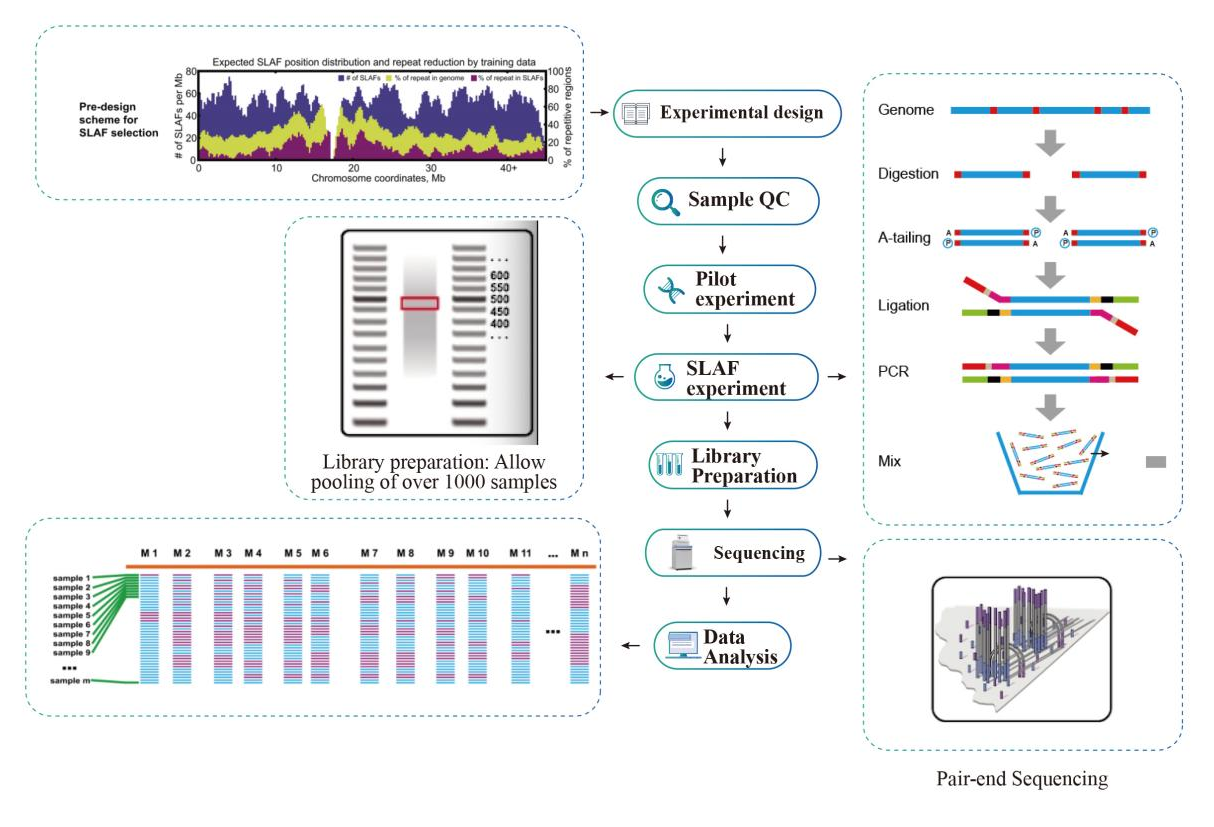
Workflow

Service fordele
Høj effektivitet til opdagelse af markører- High-throughput sekventeringsteknologi hjælper SLAF-Seq med at opdage hundredtusindvis af tags inden for hele genomet.
Lav afhængighed af genomet- Det kan anvendes på arter enten med eller uden referencegenom.
Fleksibelt skemadesign- Single-enzym, dual-enzym, multi-enzym fordøjelse og forskellige typer enzymer, alle kan vælges for at imødekomme forskellige forskningsmål eller arter.Forevaluering i silico bruges til at sikre et optimalt enzymdesign.
Effektiv enzymatisk fordøjelse- Forforsøg blev gennemført for at optimere forholdene, hvilket gør det formelle eksperiment stabilt og pålideligt.Fragmentopsamlingseffektivitet kan nå over 95 %.
Jævnt fordelt SLAF-tags- SLAF-tags er i størst udstrækning jævnt fordelt i alle kromosomer og opnår i gennemsnit 1 SLAF pr. 4 kb.
Effektiv undgåelse af gentagelser- Gentagende sekvens i SLAF-Seq-data er reduceret til lavere end 5%, især i arter med højt niveau af gentagelser, såsom hvede, majs osv.
Stor erfaring-Over 2000 lukkede SLAF-Seq-projekter på hundredvis af arter, der dækker planter, pattedyr, fugle, insekter, akvaorganismer osv.
Selvudviklet bioinformatisk arbejdsgang- En integreret bioinformatisk arbejdsgang til SLAF-Seq blev udviklet af BMKGENE for at sikre pålidelighed og nøjagtighed af det endelige output.
Service specifikationer
| Platform | Konc.(ng/gl) | I alt (ug) | OD260/280 |
| Illumina NovaSeq | >35 | >1.6(Bind>15μl) | 1,6-2,5 |
Anbefalet sekvenseringsstrategi
Sekvenseringsdybde: 10X/Tag
| Genom størrelse | Anbefalede SLAF-tags |
| < 500 Mb | 100K eller WGS |
| 500 Mb- 1 Gb | 100 K |
| 1 Gb -2 Gb | 200 K |
| Kæmpe eller komplekse genomer | 300 - 400.000 |
| Ansøgninger
| Anbefalede Befolkningsskala
| Sekvenseringsstrategi og dybde
| |
| Dybde
| Tag nummer
| ||
| GWAS
| Prøvenummer ≥ 200
| 10X
|
Ifølge genom størrelse
|
| Genetisk evolution
| Individer af hver undergruppe ≥ 10; samlede prøver ≥ 30
| 10X
| |
Anbefalet prøvelevering
Beholder: 2 ml centrifugerør
For de fleste prøver anbefaler vi ikke at konservere i ethanol.
Prøvemærkning: Prøver skal være tydeligt mærkede og identiske med den indsendte prøveinformationsformular.
Forsendelse: Tøris: Prøver skal først pakkes i poser og begraves i tøris.
Service Workflow


Prøve QC
Piloteksperiment
SLAF-eksperiment
Biblioteksforberedelse
Sekvensering
Dataanalyse
Eftersalgsservice
1. Statistik over kortresultat

2. SLAF markør udvikling

3. Variationsanmærkning

| År | Tidsskrift | IF | Titel | Ansøgninger |
| 2022 | Naturkommunikation | 17.694 | Genomisk grundlag for giga-kromosomer og giga-genom af træpæon Paeonia ostii | SLAF-GWAS |
| 2015 | Ny fytolog | 7.433 | Domesticeringsfodspor forankrer genomiske regioner af agronomisk betydning i sojabønner | SLAF-GWAS |
| 2022 | Journal of Advanced Research | 12.822 | Genomomfattende kunstige introgressioner af Gossypium barbadense i G. hirsutum afslører overlegne loci for samtidig forbedring af bomuldsfiberkvalitet og -udbytte træk | SLAF-Evolutionær genetik |
| 2019 | Molekylær plante | 10,81 | Populationsgenomisk analyse og De Novo Assembly afslører oprindelsen af Weedy Ris som et evolutionært spil | SLAF-Evolutionær genetik |
| 2019 | Naturgenetik | 31.616 | Genomsekvens og genetisk diversitet af den almindelige karpe, Cyprinus carpio | SLAF-Linkage kort |
| 2014 | Naturgenetik | 25.455 | Genomet af dyrkede jordnødder giver indsigt i bælgplantekaryotyper, polyploide evolution og afgrøde-domesticering. | SLAF-Linkage kort |
| 2022 | Plantebioteknologisk tidsskrift | 9,803 | Identifikation af ST1 afslører en udvælgelse, der involverer blaffe af frømorfologi og olieindhold under sojabønnedomestisering | SLAF-Marker udvikling |
| 2022 | International Journal of Molecular Sciences | 6,208 | Identifikation og udvikling af DNA-markører for en Wheat-Leymus mollis 2Ns (2D) Disomisk kromosomsubstitution | SLAF-Marker udvikling |





