

Genètica evolutiva









Avantatges del servei
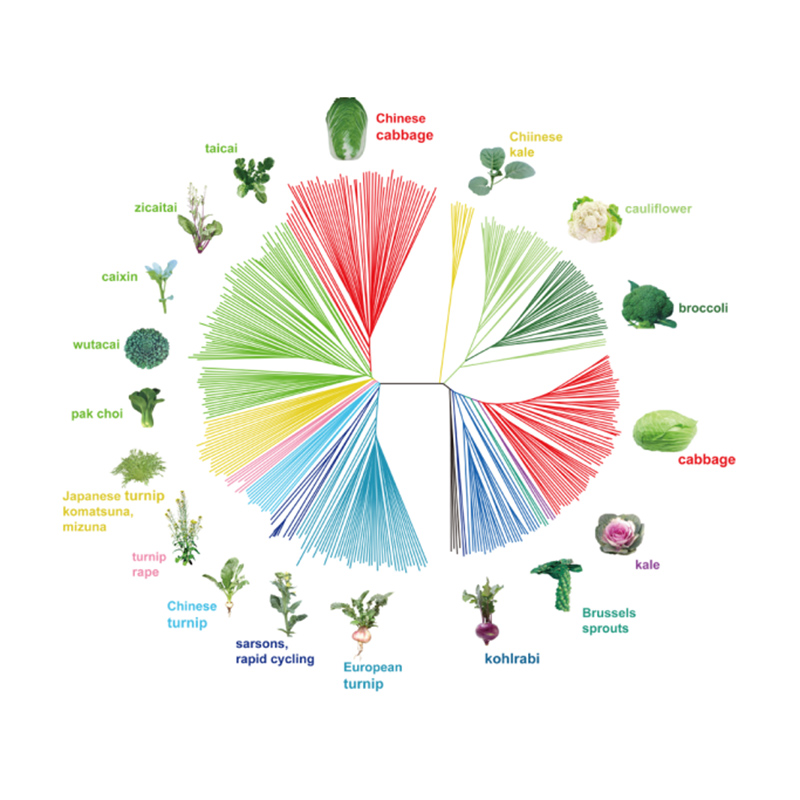
Takagi et al.,El diari de les plantes, 2013
● Estimar el temps i la velocitat de divergència de les espècies en funció de les variacions a nivell de nucleòtids i aminoàcids
● Revelació d'una relació filogenètica més fiable entre espècies amb influència minimitzada de l'evolució convergent i l'evolució paral·lela
● Construir vincles entre canvis genètics i fenotips per descobrir gens relacionats amb trets
● Estimar la diversitat genètica, que reflecteix el potencial evolutiu de les espècies
● Temps de resposta més ràpid
● Àmplia experiència: BMK ha acumulat una gran experiència en projectes relacionats amb la població i l'evolució durant més de 12 anys, abastant centenars d'espècies, etc. i ha contribuït en més de 80 projectes d'alt nivell publicats a Nature Communications, Molecular Plants, Plant Biotechnology Journal, etc.
Especificacions del servei
Materials:
Normalment, es recomana almenys tres subpoblacions (per exemple, subespècies o soques).Cada subpoblació hauria de contenir no menys de 10 individus (plantes > 15, es poden reduir per a espècies rares).
Estratègia de seqüenciació:
* WGS es pot utilitzar per a espècies amb genoma de referència d'alta qualitat, mentre que SLAF-Seq és aplicable a espècies amb o sense genoma de referència, o genoma de referència de mala qualitat.
| Aplicable a la mida del genoma | WGS | SLAF-Etiquetes (×10.000) |
| ≤ 500 Mb | 10×/individual | WGS és més recomanable |
| 500 Mb - 1 Gb | 10 | |
| 1 Gb - 2 Gb | 20 | |
| ≥2 Gb | 30 |
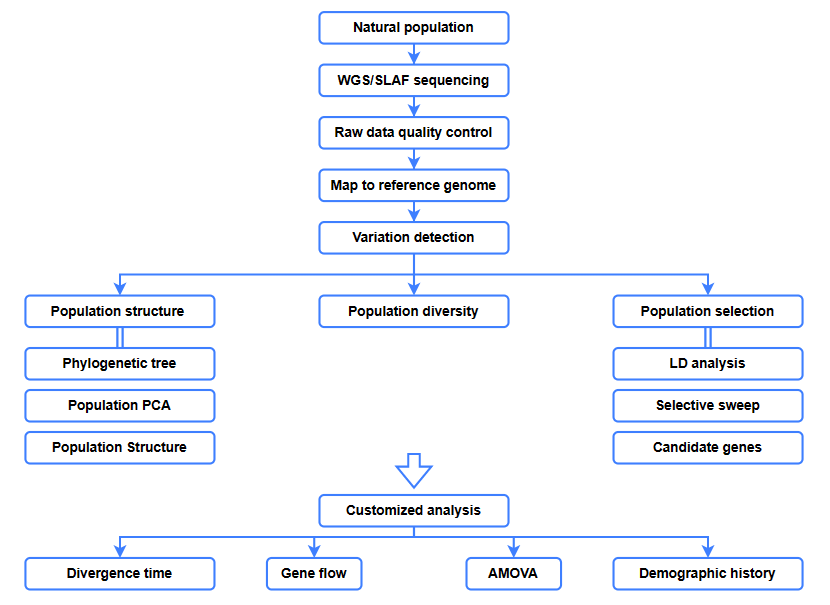
Anàlisis bioinformàtics
● Anàlisi evolutiva
● Escombrat selectiu
● Flux de gens
● Història demogràfica
● Temps de divergència
Requisits de mostra i lliurament
Requisits de mostra:
| Espècie | Teixit | WGS-NGS | SLAF |
| Animal
| Teixit visceral |
0,5 ~ 1 g
|
0,5 g
|
| Teixit muscular | |||
| Sang de mamífers | 1,5 ml
| 1,5 ml
| |
| Sang d'au/peix | |||
| Planta
| Fulla Fresca | 1 ~ 2 g | 0,5 ~ 1 g |
| Pètal/Tija | |||
| Arrel/llavor | |||
| Cèl · lules | Cèl·lula cultivada |
| gDNA | Concentració | Import (ug) | OD260/OD280 |
| SLAF | ≥35 | ≥1,6 | 1,6-2,5 |
| WGS-NGS | ≥1 | ≥0,1 | - |
Flux de treball del servei

Disseny d'experiments
Lliurament de la mostra
Construcció de la biblioteca
Seqüenciació

Anàlisi de dades
Serveis postvenda
*Els resultats de demostració que es mostren aquí són tots de genomes publicats amb BMKGENE
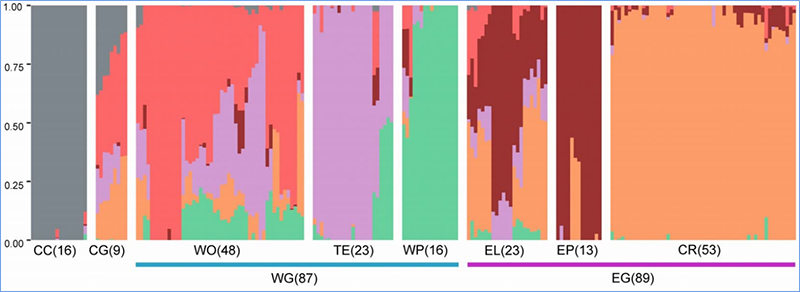
1.L'anàlisi de l'evolució conté la construcció de l'arbre filogenètic, l'estructura de la població i la PCA basada en variacions genètiques.
L'arbre filogenètic representa relacions taxonòmices i evolutives entre espècies amb avantpassat comú.
El PCA pretén visualitzar la proximitat entre subpoblacions.
L'estructura de la població mostra la presència de subpoblacions genèticament diferents en termes de freqüències d'al·lels.
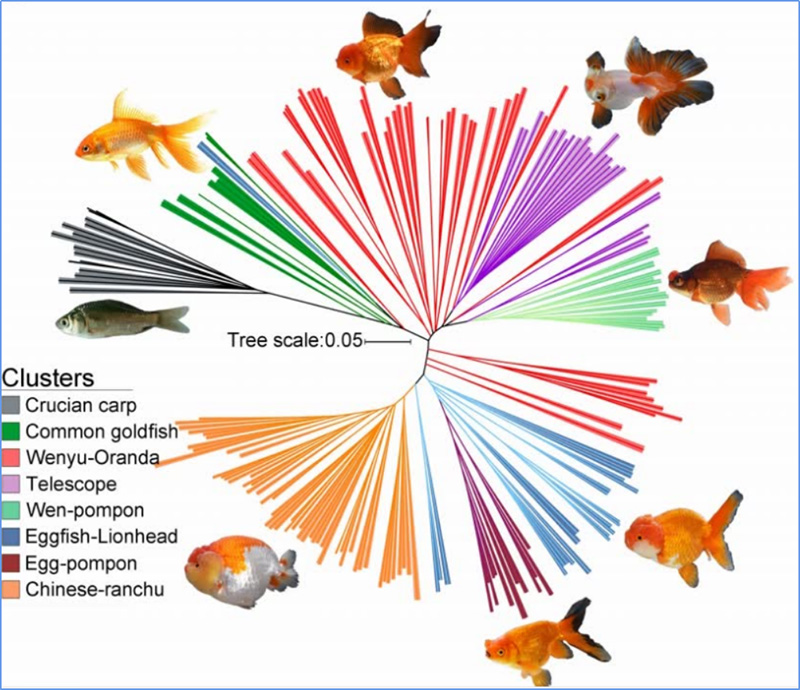
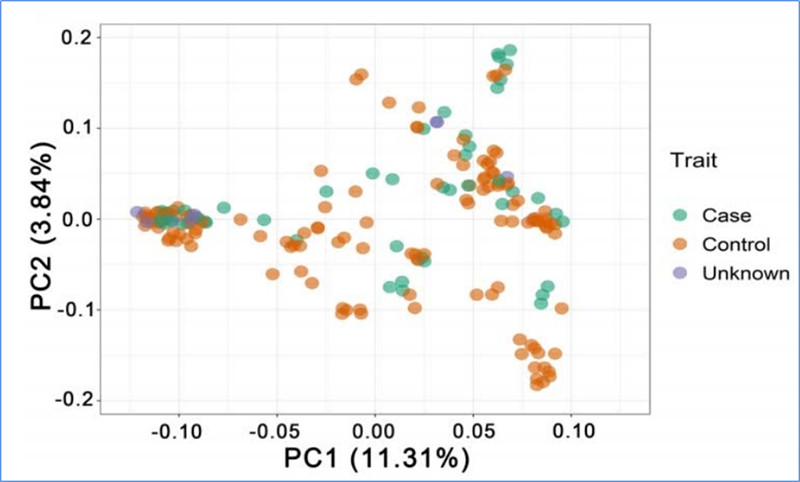
Chen, et.al.,PNAS, 2020
2. Escombrat selectiu
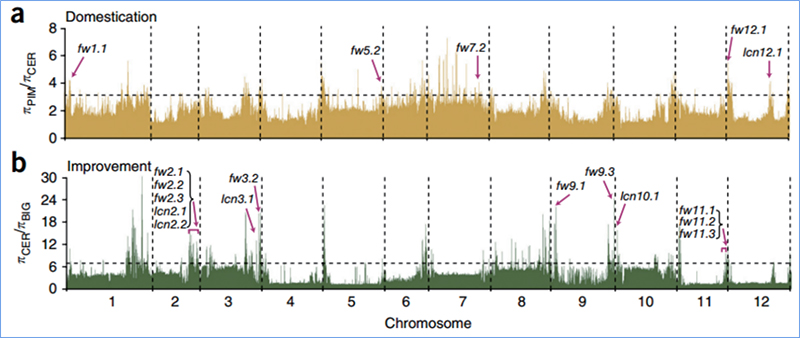
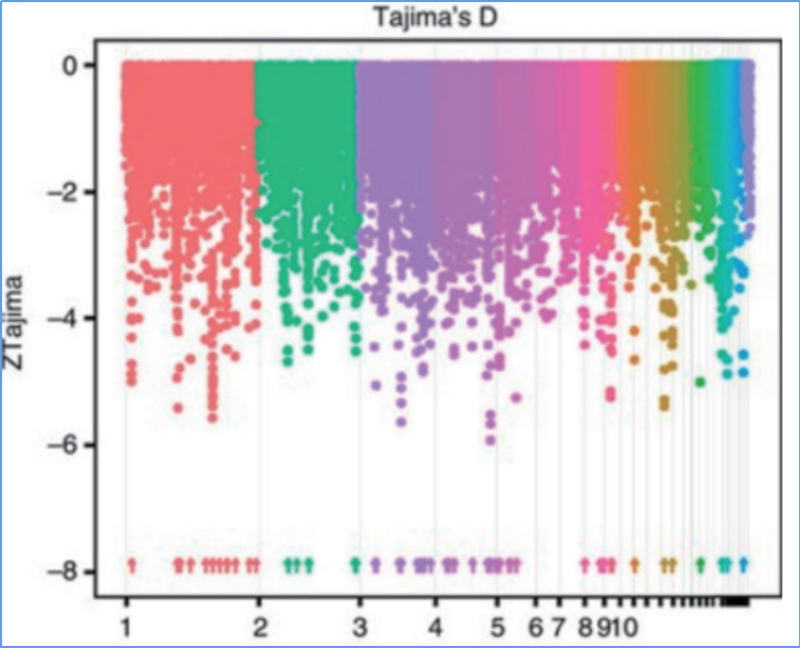
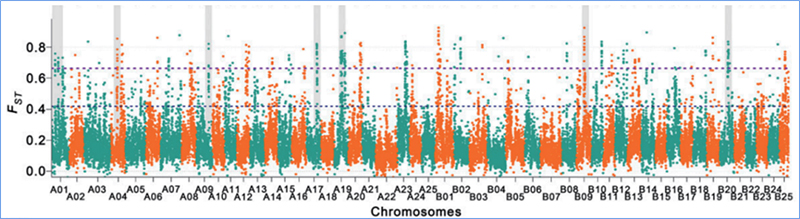
L'escombrat selectiu es refereix a un procés pel qual es selecciona un lloc avantatjós i s'augmenten les freqüències de llocs neutres enllaçats i es redueixen les dels llocs no enllaçats, la qual cosa resulta en una reducció de les regionals.
La detecció de tot el genoma a les regions d'escombrat selectiu es processa calculant l'índex genètic de la població (π,Fst, D de Tajima) de tots els SNP dins d'una finestra lliscant (100 Kb) en un pas determinat (10 Kb).
Diversitat de nucleòtids (π)
Tajima's D
Índex de fixació (Fst)
Wu, et.al.,Planta Molecular, 2018
3.Flux genètic
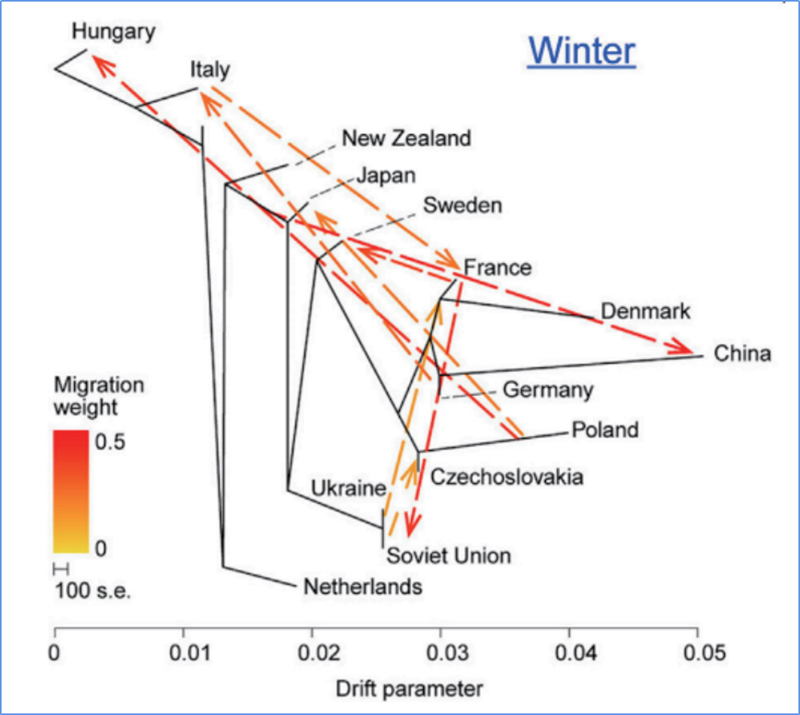
Wu, et.al.,Planta Molecular, 2018
4.Història demogràfica
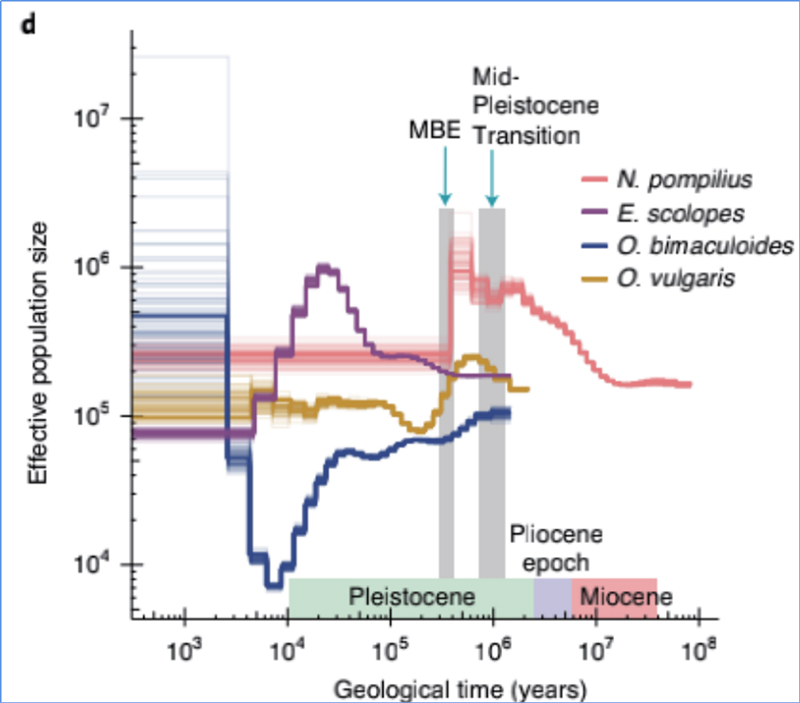
Zhang, et.al.,Natura Ecologia i evolució, 2021
5.Temps de divergència
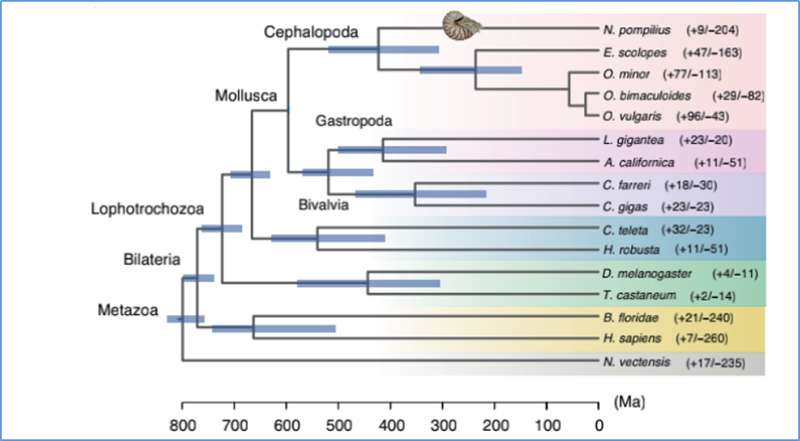
Zhang, et.al.,Natura Ecologia i evolució, 2021
Cas BMK
Un mapa de variacions genòmiques proporciona informació sobre la base genètica de la selecció de col xinesa primavera (Brassica rapa ssp. Pekinensis)
Publicat: Planta Molecular, 2018
Estratègia de seqüenciació:
Reseqüenciació: profunditat de seqüenciació: 10×
Resultats clau
En aquest estudi, es van processar 194 cols xineses per tornar a seqüenciar amb una profunditat mitjana de 10 ×, que va produir 1.208.499 SNP i 416.070 InDels.L'anàlisi filogenètica d'aquestes 194 línies va demostrar que aquestes línies es poden dividir en tres ecotips, primavera, estiu i tardor.A més, l'estructura de la població i l'anàlisi PCA van indicar que la col xinesa de primavera es va originar a partir d'una col de tardor a Shandong, Xina.Posteriorment es van introduir a Corea i el Japó, es van creuar amb les línies locals i algunes varietats d'embolcall tardà es van introduir de nou a la Xina i finalment es van convertir en col xinesa de primavera.
L'exploració de tot el genoma de les cols xineses de primavera i les cols de tardor a la selecció va revelar 23 loci genòmics que han passat per una selecció forta, dos dels quals es van solapar amb la regió de control del temps de bolting basat en el mapeig QTL.Es va trobar que aquestes dues regions contenien gens clau que regulen la floració, BrVIN3.1 i BrFLC1.Es va confirmar, a més, que aquests dos gens estaven implicats en el temps d'amortització mitjançant l'estudi del transcriptoma i els experiments transgènics.
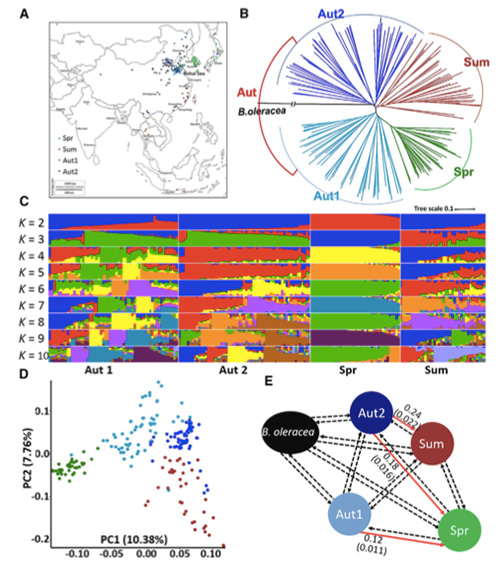 Anàlisi de l'estructura de la població de cols xineses | 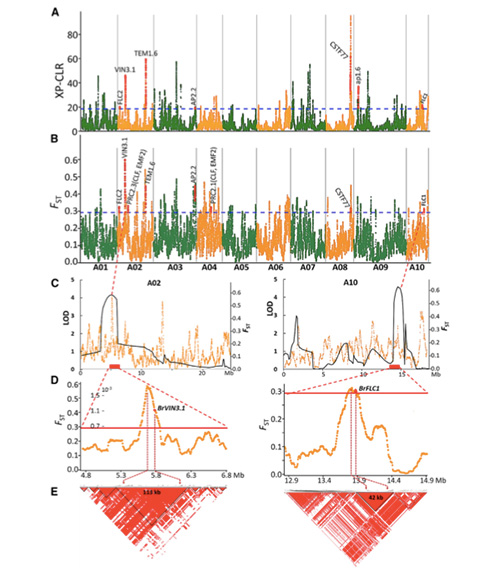 Informació genètica sobre la selecció de col xinesa |
Tongbing, et al."Un mapa de variacions genòmiques proporciona informació sobre les bases genètiques de la selecció de col xinesa de primavera (Brassica rapa ssp.pekinensis)".plantes moleculars,11 (2018): 1360-1376.





