

Еволюционна генетика









Предимства на услугата
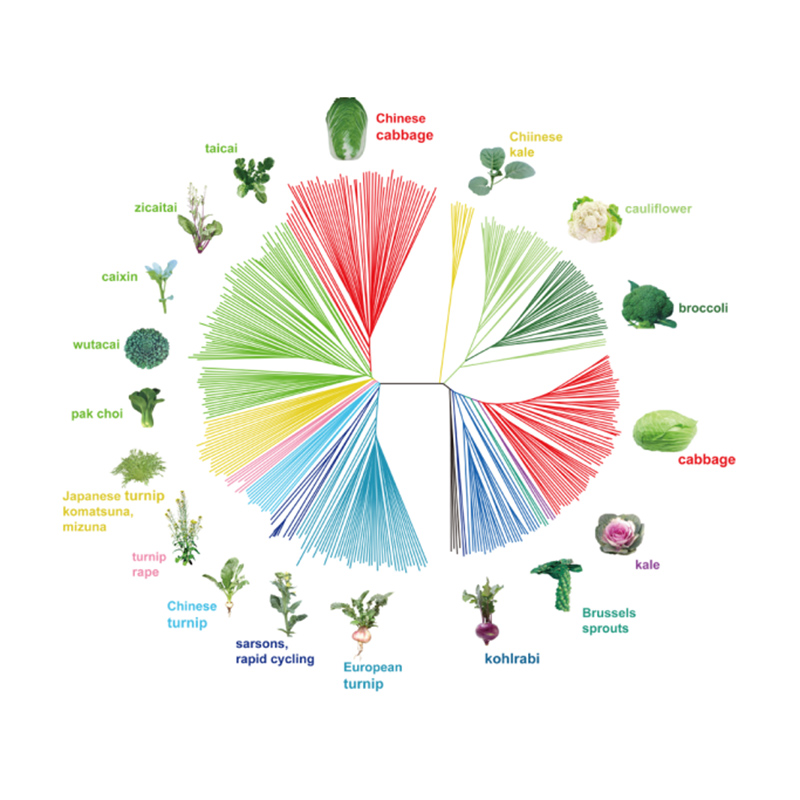
Такаги и др.,Вестникът на растенията, 2013
● Оценка на времето и скоростта на дивергенцията на видовете въз основа на вариации на ниво нуклеотиди и аминокиселини
● Разкриване на по-надеждна филогенетична връзка между видовете с минимизирано влияние на конвергентна еволюция и паралелна еволюция
● Изграждане на връзки между генетични промени и фенотипове за разкриване на гени, свързани с черти
● Оценка на генетичното разнообразие, което отразява еволюционния потенциал на видовете
● По-бързо време за изпълнение
● Обширен опит: BMK е натрупал огромен опит в популационни и еволюционни проекти за повече от 12 години, обхващащи стотици видове и т.н. и е допринесъл в над 80 проекта на високо ниво, публикувани в Nature Communications, Molecular Plants, Plant Biotechnology Journal и др.
Сервизни спецификации
Материали:
Обикновено се препоръчват поне три субпопулации (напр. подвидове или щамове).Всяка субпопулация трябва да съдържа не по-малко от 10 индивида (растения >15, може да бъде намалено за редки видове).
Стратегия за последователност:
* WGS може да се използва за видове с висококачествен референтен геном, докато SLAF-Seq е приложим за видове със или без референтен геном или референтен геном с лошо качество.
| Приложимо за размера на генома | WGS | SLAF-тагове (×10 000) |
| ≤ 500 Mb | 10×/индивидуално | WGS е по-препоръчителен |
| 500 Mb - 1 Gb | 10 | |
| 1 Gb - 2 Gb | 20 | |
| ≥2 Gb | 30 |
Биоинформатични анализи
● Еволюционен анализ
● Селективно почистване
● Генен поток
● Демографска история
● Време на отклонение
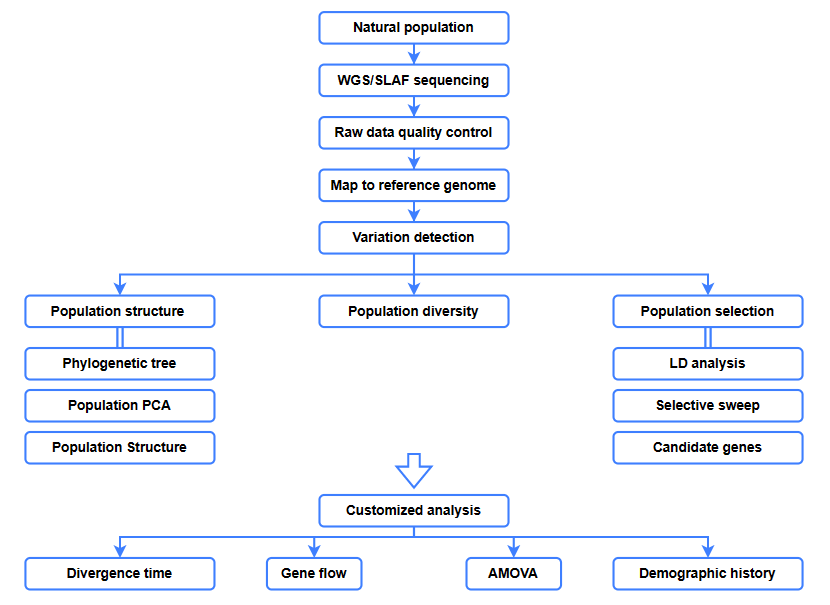
Примерни изисквания и доставка
Примерни изисквания:
| видове | Тъкан | WGS-NGS | SLAF |
| Животно
| Висцерална тъкан |
0,5~1г
|
0.5g
|
| Мускулна тъкан | |||
| Кръв на бозайник | 1,5 мл
| 1,5 мл
| |
| Кръв от птици/риба | |||
| растение
| Свежи листа | 1~2g | 0,5~1г |
| Венчелистче/Стъбло | |||
| Корен/семе | |||
| клетки | Култивирана клетка |
| gDNA | Концентрация | Количество (ug) | OD260/OD280 |
| SLAF | ≥35 | ≥1,6 | 1,6-2,5 |
| WGS-NGS | ≥1 | ≥0,1 | - |
Работен поток на услугата

Дизайн на експеримента
Доставка на мостри
Изграждане на библиотека
Секвениране

Анализ на данни
Следпродажбени услуги
*Показаните тук демонстрационни резултати са от геноми, публикувани с BMKGENE
1. Еволюционният анализ съдържа изграждане на филогенетично дърво, структура на популацията и PCA въз основа на генетични вариации.
Филогенетичното дърво представлява таксономични и еволюционни връзки между видовете с общ прародител.
PCA има за цел да визуализира близостта между подпопулациите.
Структурата на популацията показва наличието на генетично различна субпопулация по отношение на честотите на алелите.
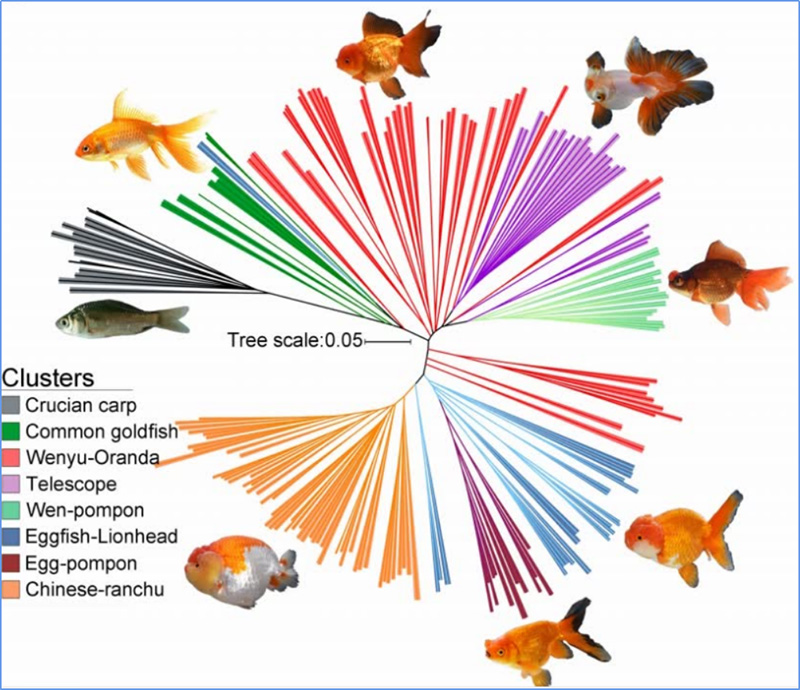
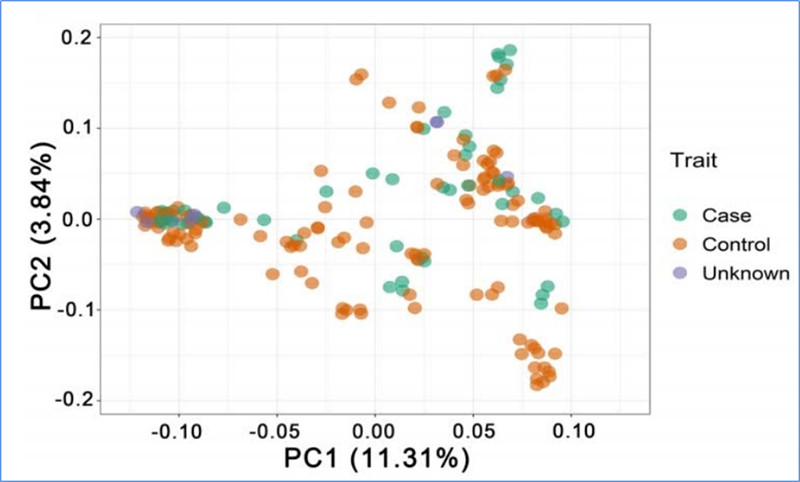
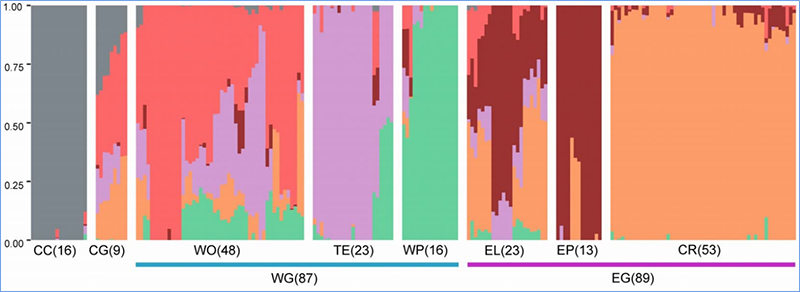
Chen, et.ал.,PNAS, 2020 г
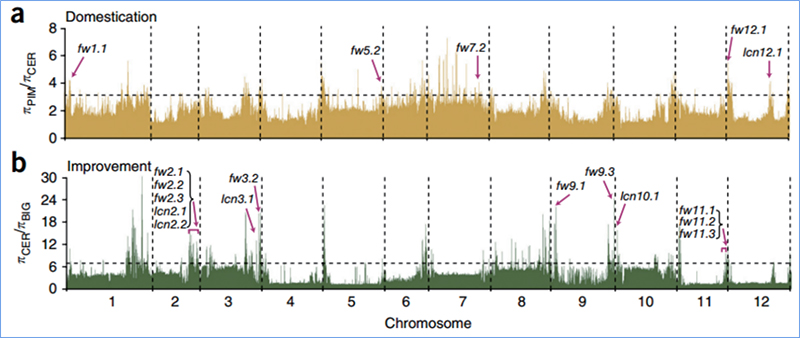
2. Селективно почистване
Селективното сканиране се отнася до процес, чрез който се избира изгоден сайт и честотите на свързаните неутрални сайтове се увеличават, а тези на несвързаните сайтове се намаляват, което води до намаляване на регионалните.
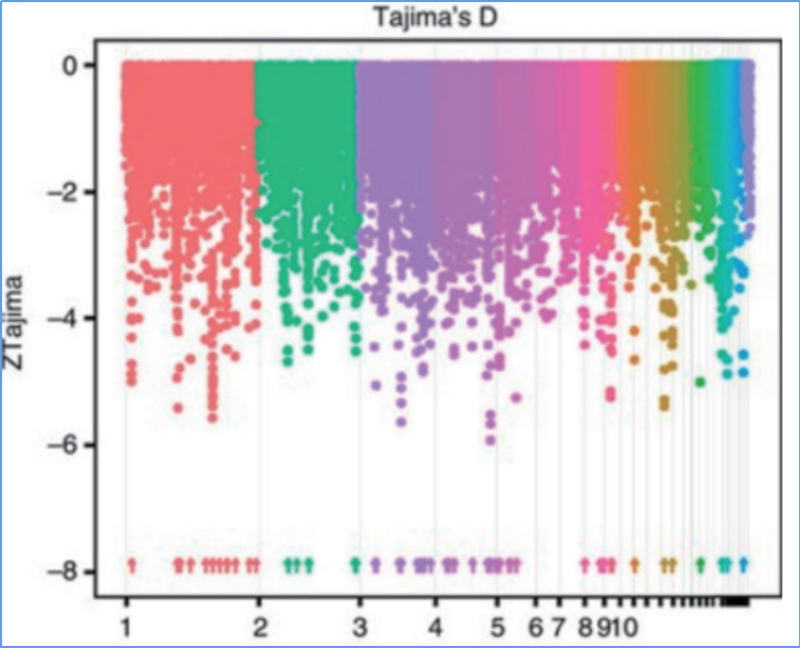
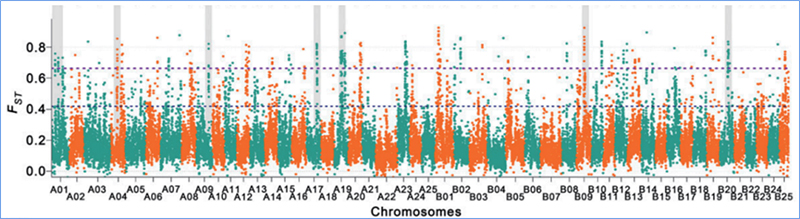
Откриването в целия геном на селективни региони за почистване се обработва чрез изчисляване на популационния генетичен индекс (π, Fst, D на Tajima) на всички SNP в рамките на плъзгащ се прозорец (100 Kb) на определена стъпка (10 Kb).
Нуклеотидно разнообразие (π)
Д. на Таджима
Индекс на фиксация (Fst)
Wu, et.ал.,Молекулярно растение, 2018 г
3. Генен поток
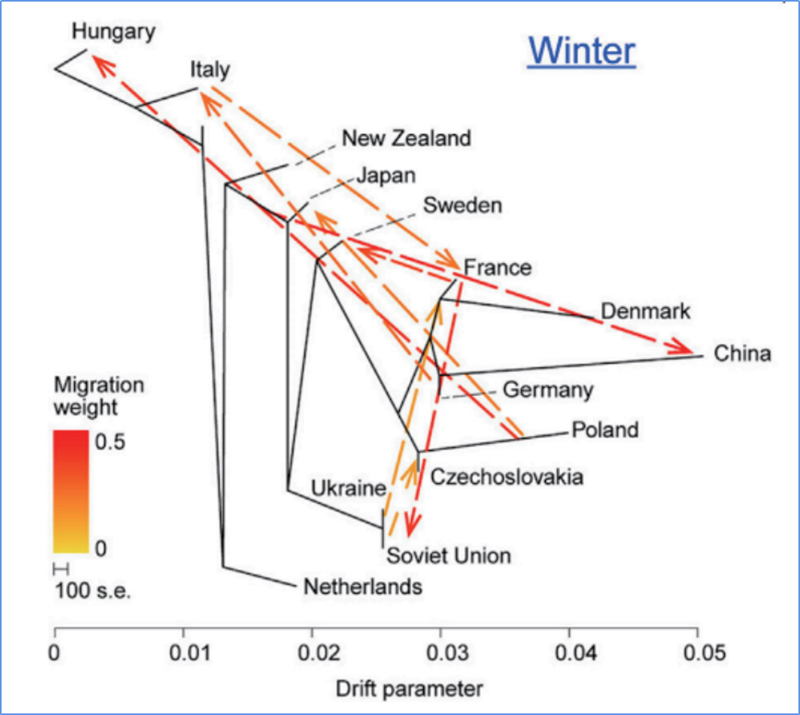
Wu, et.ал.,Молекулярно растение, 2018 г
4.Демографска история
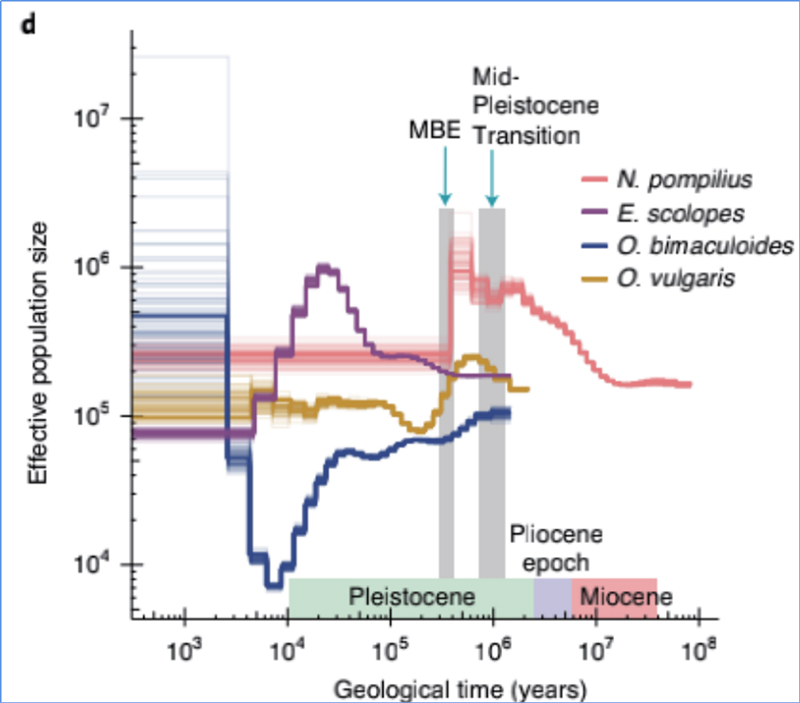
Zhang, et.ал.,Природна екология и еволюция, 2021 г
5. Време на разминаване
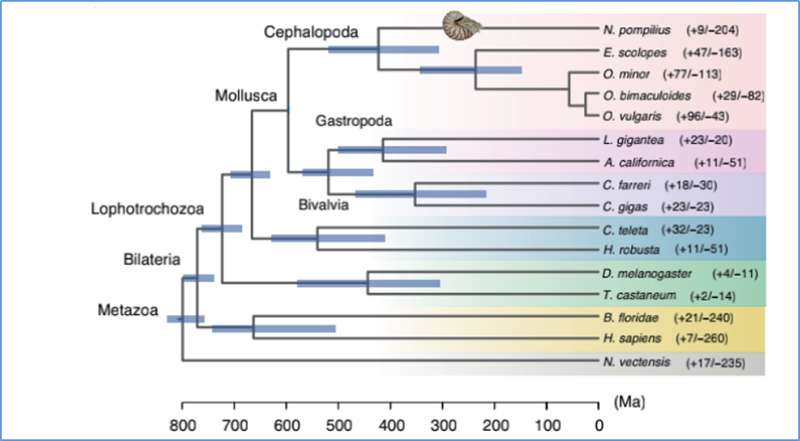
Zhang, et.ал.,Природна екология и еволюция, 2021 г
Калъф BMK
Карта на геномни вариации дава представа за генетичната основа на пролетното китайско зеле (Brassica rapa ssp. Pekinensis)
Публикувано: Молекулярно растение, 2018 г
Стратегия за последователност:
Повторно секвениране: дълбочина на секвениране: 10×
Ключови резултати
В това проучване 194 китайски зеле са обработени за повторно секвениране със средна дълбочина 10 ×, което дава 1 208 499 SNPs и 416 070 InDels.Филогенетичният анализ на тези 194 линии показва, че тези линии могат да бъдат разделени на три екотипа, пролет, лято и есен.В допълнение, структурата на популацията и PCA анализът показват, че пролетното китайско зеле произлиза от есенно зеле в Шандонг, Китай.Впоследствие те бяха въведени в Корея и Япония, кръстосани с местни линии, а някои късно закрепени разновидности от тях бяха въведени обратно в Китай и накрая станаха пролетно китайско зеле.
Общогеномното сканиране на пролетно китайско зеле и есенно зеле при селекция разкри 23 геномни локуса, които са преминали през силна селекция, два от които се припокриват с контролиращ регион на времето за болтове, базиран на QTL-картографиране.Установено е, че тези два региона съдържат ключови гени, които регулират цъфтежа, BrVIN3.1 и BrFLC1.Тези два гена бяха допълнително потвърдени, че участват във времето за завъртане чрез изследване на транскриптоми и трансгенни експерименти.
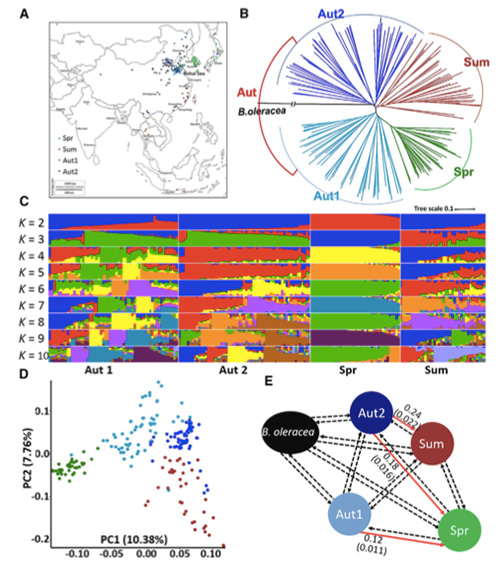 Анализ на популационната структура на китайското зеле | 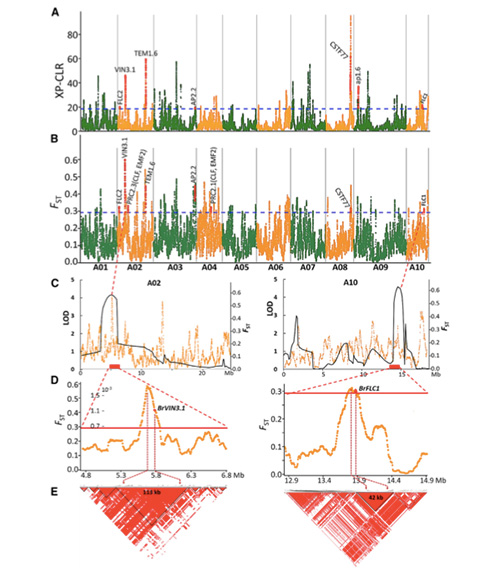 Генетична информация за селекцията на китайско зеле |
Tongbing и др.„Карта на геномните вариации дава представа за генетичната основа на селекцията от пролетно китайско зеле (Brassica rapa ssp.pekinensis).“Молекулярни растения,11 (2018): 1360-1376.





