

ЭВАЛЮЦЫЯ ГЕНОМА
генетыка прыроды
Высакаякасная зборка геному падкрэслівае геномныя характарыстыкі жыта і важныя з аграномічнага пункту гледжання гены
PacBio |Ілюміна |Аптычная карта Bionano |Зборка геному Hi-C |Генетычная карта |Выбарачныя зачысткі |РНК-Seq |ISO-seq |SLAF-пасл
Кампанія Biomarker Technologies аказала тэхнічную падтрымку ў секвенировании Pacbio, секвенировании Hi-C і аналізе даных у гэтым даследаванні.
Асноўныя моманты
1. Быў атрыманы першы высакаякасны геном Rye на храмасомным узроўні, памер адной храмасомы якога перавышае 1 Гб.
2. У параўнанні з геномамі Tu, Aet і Hv, унікальныя нядаўнія падзеі LTR-RT назіраліся ў геноме Rye, што адказвала за павелічэнне памеру геному жыта.
3. Разыходжанне паміж жытам і дыплоіднай пшаніцай адбылося пасля аддзялення ячменю ад пшаніцы, прычым час разыходжання для двух падзей склаў прыблізна 9,6 і 15 мільёнаў гадоў.
Фасфараляванне генаў FT можа кантраляваць прыкмету ранняга каласання ў жыта.
4. Выбарачны аналіз разгорткі паказвае на магчымы ўдзел ScID1 у рэгуляванні даты рублення і яго верагодную селекцыю шляхам прыручэння ў жыта
Фон
Фон
Жыта з'яўляецца каштоўнай харчовай і кармавой культурай, важным генетычным рэсурсам для паляпшэння пшаніцы і трыцікале, незаменным матэрыялам для эфектыўных параўнальных геномных даследаванняў траў.Жыта Вейнінг, раннецветущий гатунак, які культывуецца ў Кітаі, адрозніваецца сваёй шырокай устойлівасцю як да сопкай расе, так і да паласатай іржы.Каб зразумець генетычныя і малекулярныя асновы элітных прыкмет жыта і садзейнічаць геномным і селекцыйным даследаванням жыта і роднасных культур, мы тут секвеніравалі і прааналізавалі геном жыта Вейнінга.
Дасягненні
Геном жыта
Геном Rye быў пабудаваны шляхам сумяшчэння паказанняў PacBio SMRT, секвеніравання Illumina з кароткім чытаннем, а таксама з дапамогай захопу канфармацыі храмаціну (Hi-C), генетычнага картаграфавання і аналізу BioNano.Сабраныя кантыгі (7,74 Гб) складаюць 98,47% меркаванага памеру геному (7,86 Гб), прычым 93,67% кантыгаў (7,25 Гб) прыпадае на сем храмасом.Паўтаральныя элементы складалі 90,31% сабранага геному.
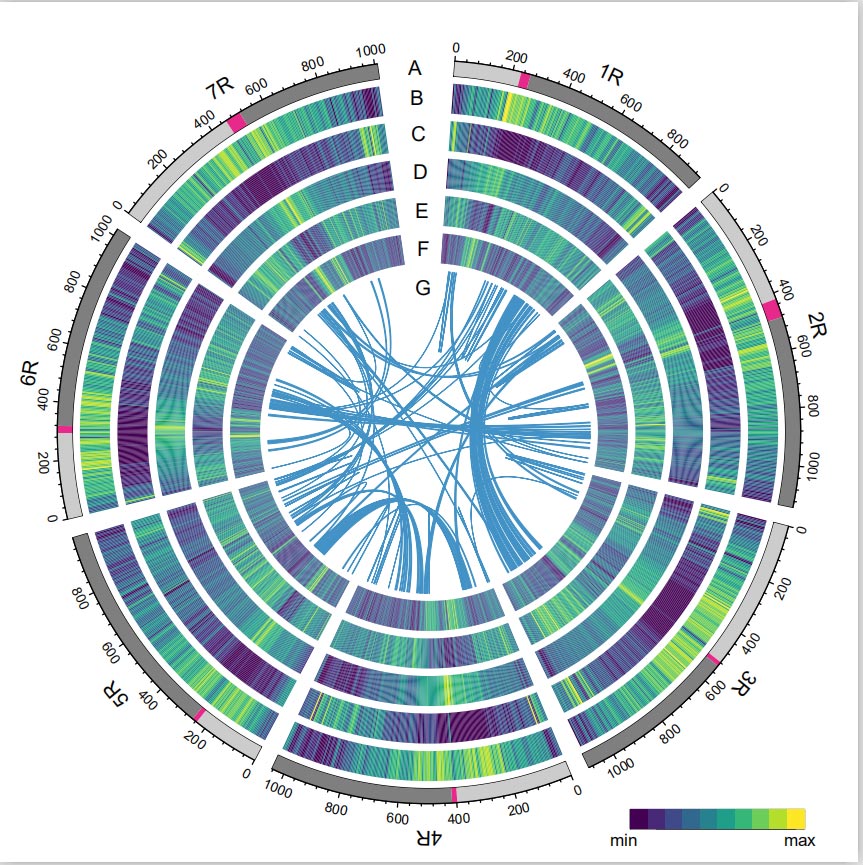
Геном жыта
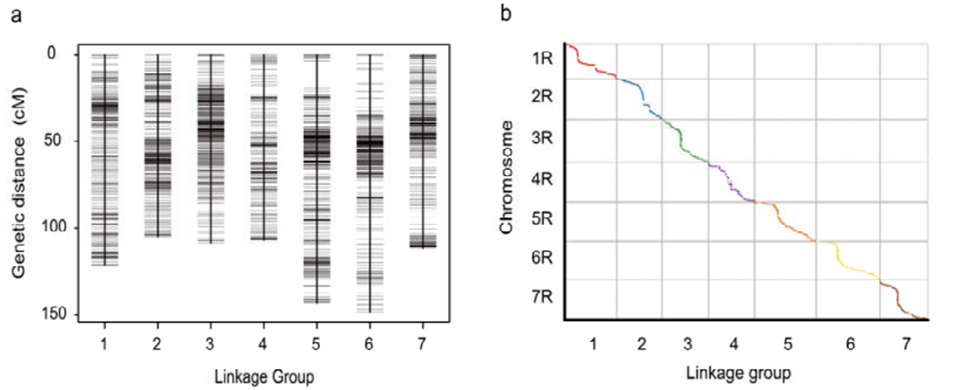
Карта генетычнай сувязі (WJ), распрацаваная з выкарыстаннем 295 раслін F2, атрыманых у выніку скрыжавання двух гатункаў жыта (Weining × Jingzhou)
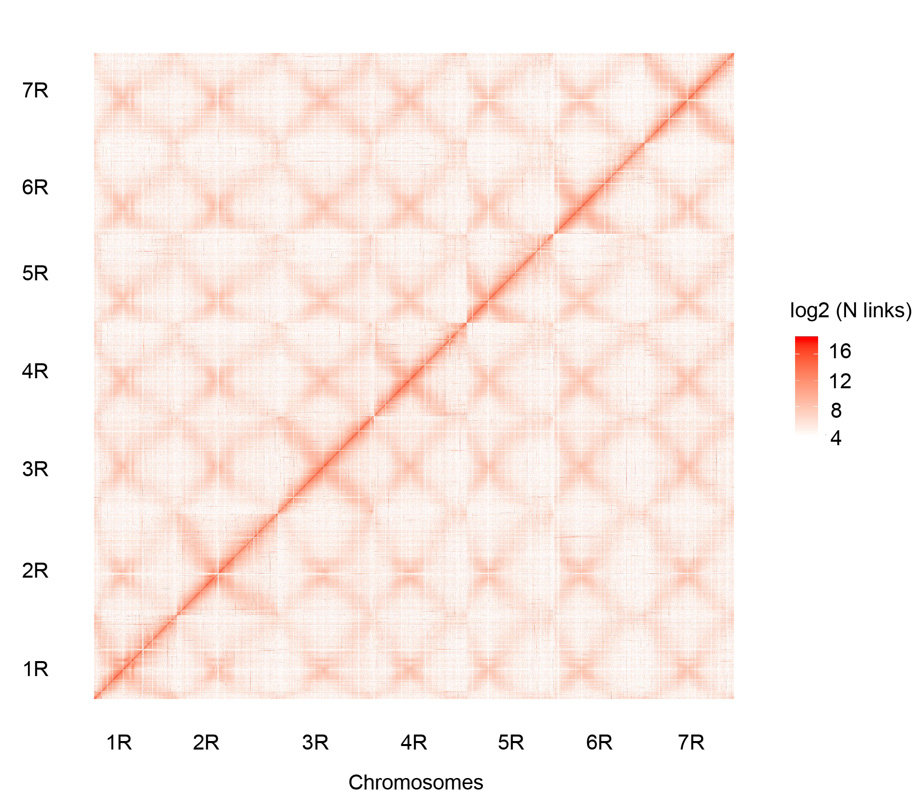
Кантактная карта Hi-C сямі сабраных жытніх храмасом Вайнінга (1R – 7R)
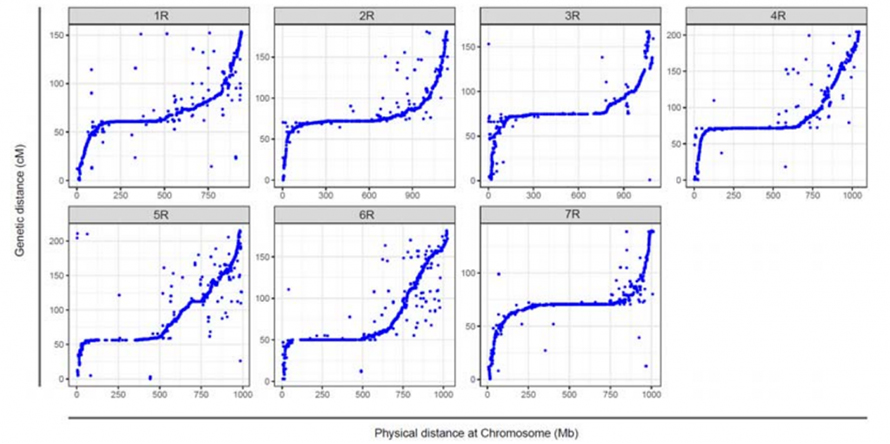
Выраўноўванне паміж сямю сабранымі храмасомамі жыта Вейнінга і сямю групамі счаплення жыта, распрацаванымі з выкарыстаннем папуляцыі RIL Lo7 x Lo255
Значэнне індэкса асамблеі LTR (LAI) геному Rye было выяўлена 18,42 і 1393 (96,74%) з 1440 высокакансерватыўных генаў BUSCO былі ідэнтыфікаваныя. Гэтыя вынікі сведчаць аб тым, што паслядоўнасць геному Weining жыта высокай якасці ў абодвух міжгенных і генетычныя рэгіёны.У агульнай складанасці было прадказана 86 991 генаў, якія кадуюць бялок, у тым ліку 45 596 генаў з высокай упэўненасцю (HC) і 41 395 генаў з нізкай упэўненасцю (LC).
2. Аналіз ТЭ
Аналіз ТЭ.У агульнай складанасці 6,99 Гб, што складае 90,31% зборкі Weining, было анатавана як TE, які ўключаў 2 671 941 элементаў, якія належаць да 537 сем'яў.Гэта ўтрыманне TE было відавочна вышэй, чым паведамлялася раней для Ta (84,70%), Tu (81,42%), Aet (84,40%), WEW (82,20%) або Hv (80,80%).Доўгія канчатковыя паўторныя рэтратранспазоны (LTR-RT), у тым ліку Gypsy, Copia і некласіфікаваныя элементы RT, былі дамінантнымі TE, і 1 займаў 84,49% анатаванага змесціва TE і 76,29% сабранага геному Вейнінга;Транспазоны CACTA ДНК былі другімі па колькасці TE, складаючы 11,68% анатаванага ўтрымання TE і 10,55% сабранага геному Вайнінга.
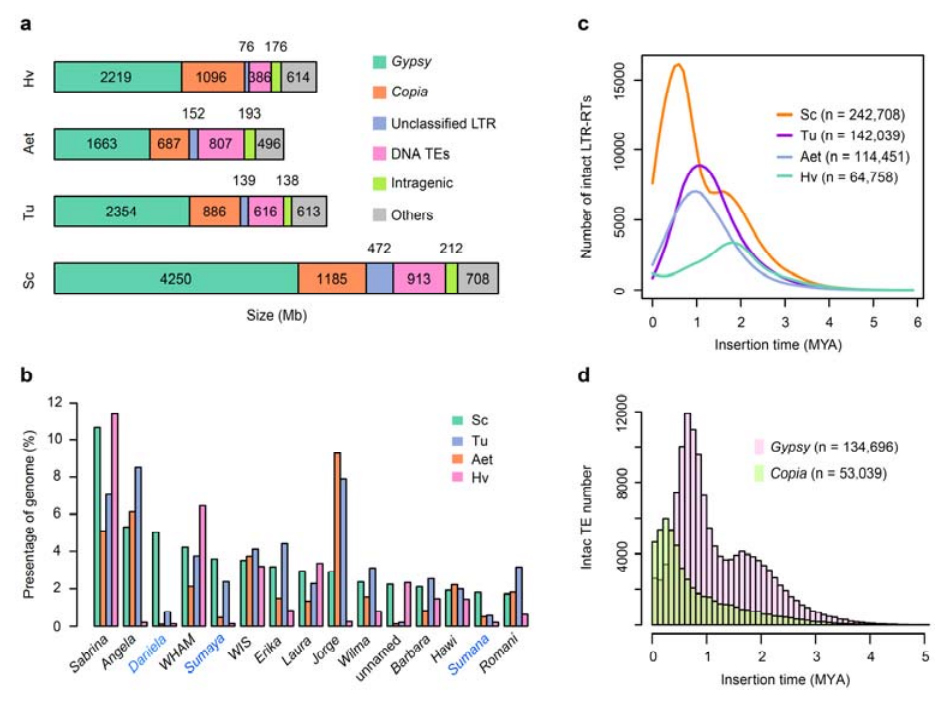
Аналіз транспозонных элементаў жыта
У жыта Вейнінга была параўнальна высокая доля нядаўніх уключэнняў LTR-RT з пікам узмацнення, які з'явіўся прыкладна 0,5 мільёна гадоў таму (MYA), што было самым апошнім сярод чатырох відаў;другі пік, які адбыўся прыблізна праз 1,7 мільёна гадоў года, быў старэйшы і таксама назіраўся ў ячменю.На ўзроўні звышсямейства былі знойдзены зусім нядаўнія ўсплёскі элементаў Copia ў жыце Weining на 0,3 мільёна гадоў года, у той час як узмацненне цыганскіх RT дамінантна фармавала бімадальны ўзор размеркавання дынамікі выбухаў LTR-RT.
3. Даследаванне эвалюцыі геному жыта і сінтэніі храмасом
Разыходжанне паміж жытам і дыплоідным пшаніцай адбылося пасля аддзялення ячменю ад пшаніцы, прычым час разыходжання для двух падзей склаў прыблізна 9,6 і 15 мільёнаў гадоў адпаведна.1R, 2R, 3R былі цалкам коллинеарны з групамі 1, 2 і 3 храмасом пшаніцы, адпаведна.4R, 5R, 6R, 7R былі выяўленыя існуе буйнамаштабныя зліцця і сегменты.
4. Аналіз дуплікацый генаў і іх уплыў на гены біясінтэзу крухмалу
Характэрна, што колькасць тандэмна дубліраваных генаў (TDG) і праксімальна дубліраваных генаў (PDG) у жыта Вейнінга была вышэй, чым у Tu, Aet, Hv, Bd і Os.Транспанаваныя дубляваныя гены (TrDG) таксама былі больш шматлікія, чым тыя, якія былі спецыяльна знойдзены для Tu і Aet.Пашырэнне геному жыта суправаджаецца большай колькасцю дубляцый генаў.Павелічэнне выбухаў TE ў жыце магло прывесці да павышэння колькасці TrDG.
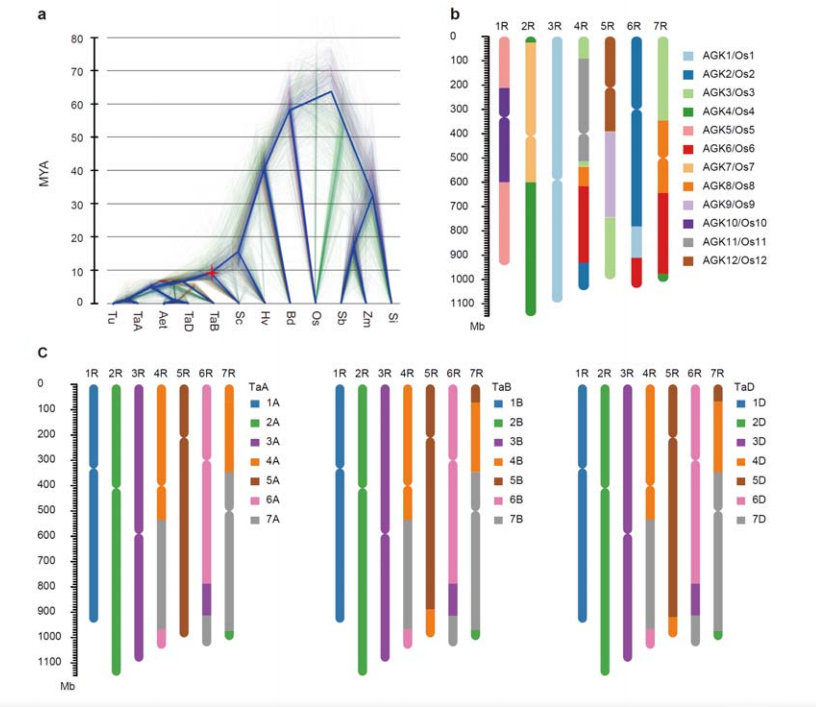
Эвалюцыйны і храмасомна-сінтэнічны аналіз геному жыта
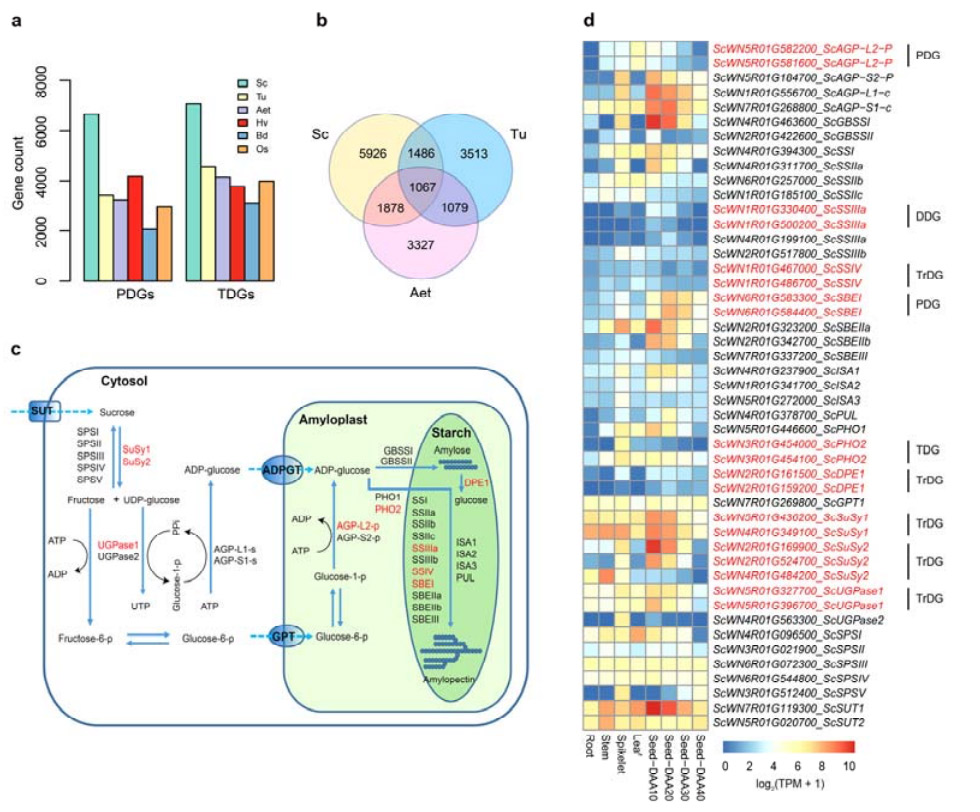
Аналіз дубліравання генаў жыта і іх уплыў на разнастайнасць генаў, звязаных з біясінтэзам крухмалу (SBRG)
5. Раскрыццё локусов гена запаснага бялку насення жыта (SSP).
Чатыры храмасомных локуса (ад Sec-1 да Sec-4), якія вызначаюць SSP жыта, былі выяўлены на 1R або 2R.Гены α-глиадина з'явіліся толькі нядаўна ў пшаніцы і блізкіх відаў пасля аддзялення пшаніцы ад жыта.
6. Даследаванне фактару транскрыпцыі (ТФ) і генаў устойлівасці да хвароб
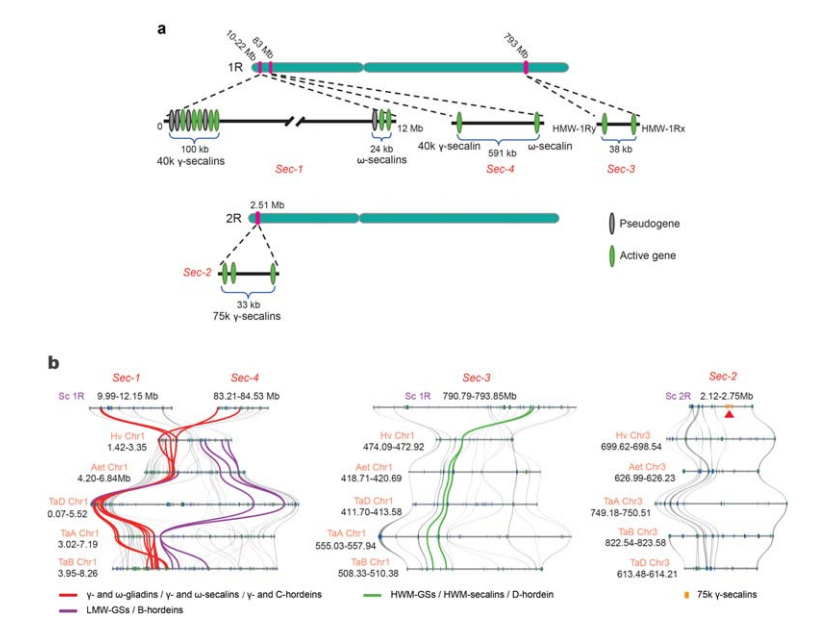
Аналіз локусов секалина жыта
У жыта Вейнінга больш генаў, звязаных з устойлівасцю да хвароб (DRA) (1989, дадатковыя дадзеныя 3), чым у Tu (1621), Aet (1758), Hv (1508), Bd (1178), Os (1575) і A (1836). ), B (1728) і D (1888) субгеномы звычайнай пшаніцы.
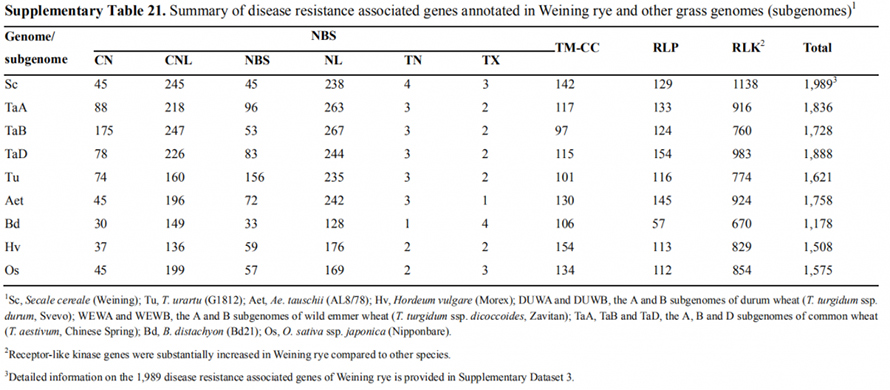
7. Даследаванне асаблівасцяў экспрэсіі генаў, звязаных з прыкметай ранняга загалоўка
Два гена FT з адносна высокай экспрэсіяй ва ўмовах доўгага дня, ScFT1 і ScFT2, былі анатаваны ў зборцы геному Вейнінга.Выяўлена ўзаемасувязь двух амінакіслот фасфаралявання ScFT2 (S76 і T132) са зніжэннем кантролю часу
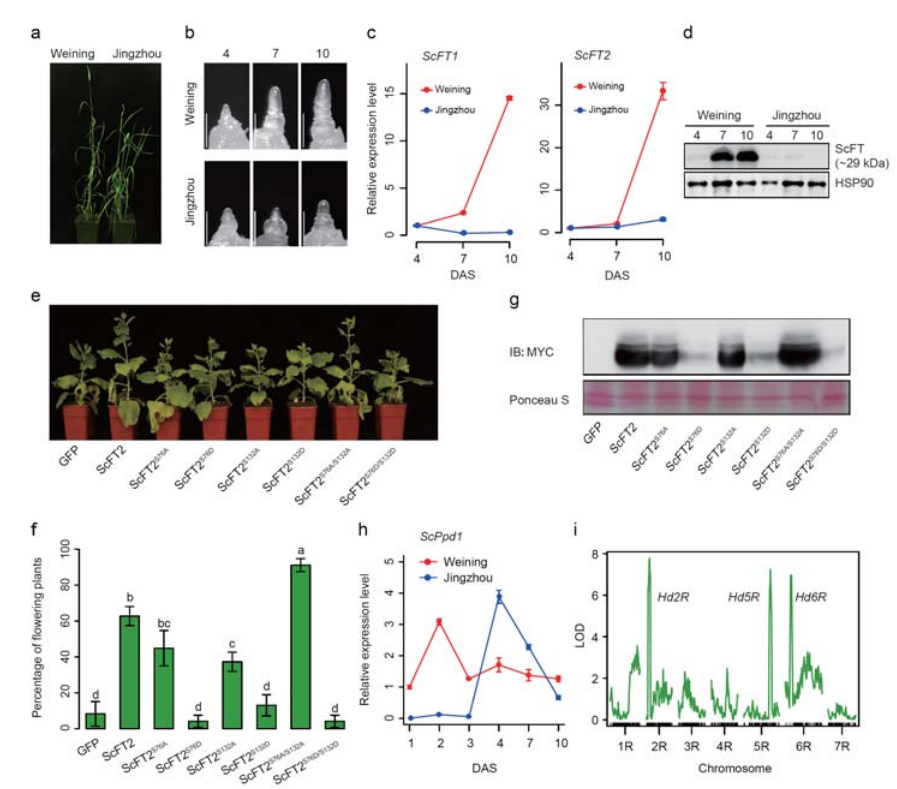
Развіццё і асаблівасці экспрэсіі генаў, звязаныя з ранняй загаловак прыкмета Weining жыта
8. Здабыча храмасомных абласцей і локусов, якія патэнцыйна ўдзельнічаюць у прыручэнні жыта
У агульнай складанасці 123 647 SNP былі выкарыстаны для правядзення селектыўнага аналізу разгорткі паміж культывуемым жытам і S. vavilovii.11 селектыўных сігналаў разгорткі, вызначаных індэксам памяншэння (DRI), індэксам фіксацыі (FST) і метадам XP-CLR.Было выяўлена магчымае ўдзел ScID1 у рэгуляванні даты загалоўка.
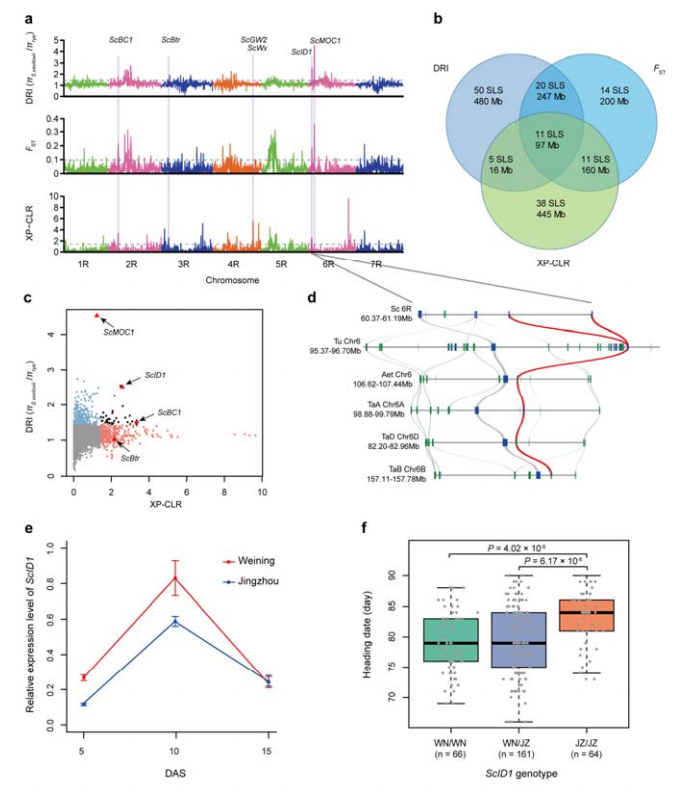
Ідэнтыфікацыя і аналіз храмасомных абласцей і локусов, патэнцыйна звязаных з прыручэннем жыта
Даведка
Li GW і інш.Высакаякасная зборка геному падкрэслівае геномныя характарыстыкі жыта і важныя з аграномічнага пункту гледжання гены.Генетыка прыроды (2021)
Навіны і асноўныя моманты накіравана на абмен апошнімі паспяховымі кейсамі з Biomarker Technologies, фіксаванне новых навуковых дасягненняў, а таксама вядомых метадаў, якія прымяняюцца падчас даследавання.
Час публікацыі: 5 студзеня 2022 г